
|
|||
|---|---|---|---|
|
|
Biogénesis mitocondrial y ejercicio |
|
|
|
Licenciada en Ciencias Biológicas con orientación en Biología Celular Becaria de Doctorado del Consejo Nacional de Investigaciones Científicas y Tecnológicas de Argentina (CONICET), en el Laboratorio de Neurobiología del Desarrollo del Instituto de Investigaciones Bioquímicas de Bahía Blanca - INIBIBB. |
Lic. María Fernanda Insua mfinsua@criba.edu.ar (Argentina) |
|
|
|
|
http://www.efdeportes.com/ Revista Digital - Buenos Aires - Año 9 - N° 61 - Junio de 2003 |
|
|
1 / 1
La actividad contráctil crónica produce biogénesis mitocondrial en el músculo, entendiendo estos términos como los procesos celulares involucrados en la síntesis y degradación de estas organelas.
Mecanismos celulares de biogénesis mitocondrialLa iniciación de la biogénesis mitocondrial en las células musculares comienza con ciertas señales provocadas por la contracción del músculo. La magnitud de la señal/es, indudablemente hasta cierto punto, está relacionada con la intensidad y duración del esfuerzo contráctil. Esta señalización puede potencialmente conducir a 1) la activación o inhibición de factores de trascripción, los cuales afectarán la velocidad de trascripción, 2) la activación o inhibición de factores de estabilización del RNAm, los cuales median cambios en la velocidad de degradación del RNAm, 3) alteraciones en la eficiencia de la traducción a proteínas, 4) la modificación post-traduccional de las proteínas, 5) cambios en la cinética de importación de proteínas mitocondriales y/o 6) alteraciones en la velocidad de ensamblado de las subunidades que conforman los complejos enzimáticos mitocondriales.
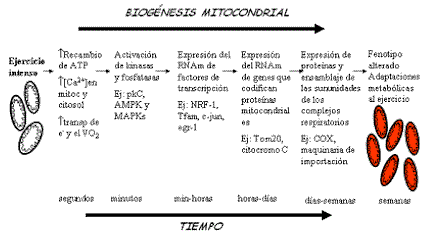
Lic. M Fernanda Insua (Modificado de Hoods, 2001)Además, la señal/es podría ser transmitida directamente a la mitocondria para iniciar la replicación o trascripción del ADNmitocondrial (ADNmt), o tener un efecto sobre la traducción del RNAm a proteínas o en el ensamblado de los complejos enzimáticos.
Eventos iniciales de señalizaciónSi hablamos de la actividad contráctil, al inicio de la misma, se sucede un número de eventos rápidos, que forman parte del proceso de señalización inicial, que conduce a la síntesis de proteínas y lípidos. Estos incluyen: 1) cambios en la conformación de proteínas sensibles al voltaje, respondiendo a potenciales de acción del sarcolema (membrana plasmática del músculo); 2) activación de unas proteínas de la superficie celular llamadas integrinas, que son mecano-transductores (traducen señales mecánicas desde el exterior al interior celular); 3) flujo de iones (por ej. de Ca2+) en la célula muscular en contracción; 4) cambios en la duración de los puentes cruzados y el desarrollo de la tensión y 5) el recambio de ATP y la subsiguiente estimulación del metabolismo.
Se discutirán principalmente los eventos relacionados con la señalización por calcio y el recambio de ATP.
Calcio. Su liberación del retículo sarcoplásmico, permite la interacción de la actina y la miosina en la célula muscular. También es muy reconocido como un importante segundo mensajero en una variedad de células, incluyendo las musculares. Incrementos en la concentración de calcio citosólico pueden activar un número de proteínas kinasas (por ej la Ca2+/calmodulina kinasa II, la proteína kinasa C (PKC)) y fosfatasas (por ej, la calcineurina), las cuales en última instancia envían sus señales al núcleo para alterar la velocidad de la trascripción de ciertos genes (proceso de síntesis de ARNm a partir del ADN).
Existen otros eventos en los que participa el calcio que podrían estar relacionados con la biogénesis mitocondrial. Primero, el incremento de calcio citosólico influencia directamente la velocidad de la respiración u oxidación mitocondrial. Esto ocurre a través de la activación de enzimas deshidrogenasas, las cuales requieren calcio para su actividad completa. El cambio en el calcio mitocondrial causa incrementos en los niveles de ATP tanto en el citosol como en la mitocondria, los cuales permanecen más tiempo, luego de que las elevaciones de calcio mitocondrial han declinado.
En segundo término, se han reportado cambios mediados por calcio en eventos de fosforilación en la membrana mitocondrial interna, particularmente con la fosforilación de la subunidad c de la ATPasa F0 F1 (la que participa en la síntesis de ATP). Aunque el significado de estos hechos respecto a la biogénesis mitocondrial aún no ha sido establecido, podrían estar formando parte de una cascada de señalización intramitocondrial.
Tercero, disminuciones en la concentración de calcio en el medio que baña miotubos en cultivo, produce cambios paralelos en las actividades de las enzimas mitocondriales. En cuarto lugar, han sido documentados incrementos mediados por calcio en la expresión de citocromo c, un importante componente de la cadena de transporte de electrones.
Finalmente, estudios realizados en células con una depleción inducida del ADN mitocondrial (ADNmt), han sostenido la conclusión de que algunos eventos señalizados por calcio en el músculo, son propagados como resultado directo de desbalances en la relación suministro-demanda de energía.
En la depleción del ADNmt, la síntesis de ATP está reducida por una cadena respiratoria defectuosa, resultado de niveles inadecuados de ciertas proteínas codificadas por el genoma mitocondrial. Esto conduce a un incremento de la concentración de calcio citosólico, presumiblemente porque los procesos dependientes de energía como la extrusión (salida) y la entrada de calcio están deteriorados.
Más recientemente Wu y col. generaron un ratón transgénico que expresa selectivamente en el músculo esquelético, una forma constitutivamente activa de la proteína kinasa IV dependiente de calcio/calmodulina (CaMKIV). Los músculos esqueléticos de estos ratones mostraron una replicación aumentada de ADN mitocondrial y biogénesis de mitocondrias, una regulación aumentada de las enzimas mitocondriales involucradas con el metabolismo de los ácidos grasos y el transporte de electrones, y una susceptibilidad reducida a la fatiga durante contracciones repetitivas. La CaMK indujo la expresión de PGC-1 (peroxisome proliferator-activated receptor gamma coactivator 1 (PGC-1), un regulador maestro de la biogénesis mitocondrial in vivo, y activó el promotor del gen PGC-1 en miocitos en cultivo. Por lo tanto una vía de señalización regulada por calcio, controla la biogénesis mitocondrial en células de mamífero (Wu, 2002).
Sin embargo, algunos estudios han mostrado que un incremento de calcio no puede, por sí mismo, conducir a un aumento de la biogénesis mitocondrial, sino que estaría formando sólo parte de una serie más amplia de señales positivas y negativas que median los cambios en la síntesis de componentes mitocondriales.
Recambio de ATPPor muchos años, ha sido hipotetizado que disturbios en el metabolismo energético que conduce a una depleción de ATP, una alteración en la carga energética, o un cambio en el potencial de fosforilación, podrían iniciar una respuesta compensatoria, conduciendo en última instancia a un incremento del contenido mitocondrial. Sin embargo, el entrenamiento físico de moderada intensidad puede conducir a la biogénesis mitocondrial en ausencia de cambios notables en los niveles celulares de ATP. Por lo tanto, a pesar de que las condiciones de depleción de ATP son conocidas por conducir a un contenido mayor de mitocondrias, pareciera ser que un incremento en la velocidad de recambio del ATP (es decir, velocidades aumentadas de síntesis y degradación de ATP), es suficiente para provocar la biogénesis mitocondrial. Estas ideas se apoyan en un considerable soporte experimental: experimentos realizados con una droga (el ácido b-guanidinopropriónico) para depletar los compuestos con grupos fosfatos de alta energía como la fosfocreatina y el ATP, muestran una regulación elevada de las enzimas mitocondriales y el ARNm del citocromo c, en músculo. En segundo término, como resultado del recambio acelerado de ATP, el ejercicio incrementa los niveles de AMP y disminuye los de fosfocreatina en el músculo, provocando la activación de una proteína kinasa, la a2-AMPK (ver fig. 1).
El desacople o desacoplamiento mitocondrial se refiere a la disipación del gradiente electroquímico a través de la membrana interna, lo cual disminuye la producción de ATP, inhibiendo la actividad de la ATPasa F1 F0. Subsecuentemente, el transporte de electrones y el consumo de oxígeno se aceleran porque se pierde el control ejercido por el ADP, y la utilización de ATP excede la síntesis. En este sentido, esta condición es análoga al ejercicio intenso.
El desacople puede lograrse por el uso de drogas específicas in vitro, o por la sobreexpresión de proteínas de desacople (UCPs). En 6 hs. de desacople, se observó una inducción del factor de trascripción llamado factor-1 respiratorio nuclear (NRF-1). NRF-1 ha sido ampliamente involucrado en la activación de múltiples genes implicados en la biogénesis mitocondrial. Seguidamente a la inducción del NRF-1, se observó un aumento en la expresión de los genes que son target o blanco de este factor, incluyendo la d-ácido amino levulínico sintasa (ALAs), la enzima limitante de la velocidad en la síntesis del grupo hemo. Este grupo es un componente vital de la cadena respiratoria, ya que actúa como grupo prostético de los citocromos mitocondriales. Por lo tanto parece que el incremento de la respiración mitocondrial, o el déficit entre la demanda celular de ATP y la provisión de ATP mitocondrial, proporciona un estímulo para la subsecuente inducción de una variedad de genes involucrados en la biogénesis de la organela.
En relación a las proteínas de desacople, la función fisiológica de la proteína de desacople-3 (UCP3) aún no ha sido dilucidada. Estudios previos muestran que el ejercicio induce aumentos muy considerables de la proteína UCP3 en el músculo esquelético. Como la expresión de UCP3 parece estar regulada por los mismos mecanismos que rigen otros componentes mitocondriales, resultaba poco probable que el ejercicio produjera esos cambios tan impresionantes y divergentes en la composición mitocondrial.
Jones y col, testearon la hipótesis de que los principales cambios en la concentración de proteína UCP3 no ocurren independientemente de la biogénesis mitocondrial, y que UCP3 aumenta como parte del incremento de mitocondrias inducido por el ejercicio.
Ellos encontraron un gran aumento del ARNm de UCP3 inmediatamente y 3 hs. después de una sesión de natación. La concentración de proteína UCP3 se incrementó aproximadamente 35% 18hs después de una única sesión de ejercicio, aproximadamente 63% luego de 3 días, y aproximadamente 84% luego de 10 días. Estos resultados son consistentes con la interpretación acerca de que el ejercicio de resistencia induce un incremento adaptativo en la mitocondria que posee un contenido normal de UCP3. Esta proteína también aumenta durante el ayuno y las comidas altas en grasa. En estas situaciones la disponibilidad de ácidos grasos excede la capacidad de metabolizarlos, sugiriendo que la UCP3 cumpliría un rol de regulación y manejo de estos ácidos grasos no metabolizables. Para testear la hipótesis que indica que esta proteína actuaría como exportadora de ácidos grasos no metabolizables desde la matriz mitocondrial, Schrauwen y col., le suministraron a pacientes Etomoxir (bloqueador de la entrada de ácidos grasos a la mitocondria), o un placebo, durante 36hs en una cámara respiratoria. El etomoxir inhibió la oxidación de grasa durante 24hs y la oxidación de grasas durante el ejercicio aproximadamente entre 14-19%. Sorpresivamente, el contenido de UCP3 en el músculo vastus lateralis, sufrió una marcada up-regulación en las 36hs de la administración de Etomoxir. Este aumento de la UCP3 fue acompañado por una glucosa sanguínea en ayuno más baja, y un incremento de la traslocación del transportador de glucosa 4 (GLUT-4).
Estos datos apoyan la hipótesis que supone que la función fisiológica de la UCP3 es facilitar la salida de los ácidos grasos no metabolizables desde la matriz mitocondrial hacia el citosol, lo cuál preserva a la mitocondria de los potenciales efectos deletéreos de niveles altos de ácidos grasos. Además los datos muestran que la regulación aumentada de la UCP3, puede ser beneficiosa en el tratamiento de la diabetes tipo2 (Schrauwen, 2002).
En conclusión, existe fuerte evidencia para sostener roles interactivos y complementarios de la señalización por calcio y por disturbios en el recambio de ATP, como importantes contribuyentes a la iniciación de los eventos que conducen a la biogénesis mitocondrial.
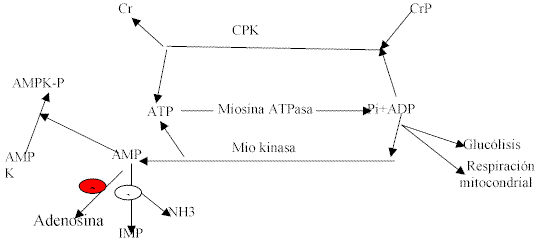
Fig. 1: metabolismo del nucleótido adenina durante el ejercicio intenso. Desde el inicio hasta la finalización de las contracciones, el ADP libre (ADPl) se forma por la reacción de la miosina ATPasa. El ADPl se combina con la fosfocreatina (CrP) para dar creatina, mediante la reacción de la creatina fosfokinasa (CPK), provocando niveles reducidos de CrP y restauración de ATP. El ADPl también estimula la glucólisis y la respiración mitocondrial. 2 moléculas de ADPl pueden ser convertidas a ATP y AMP en la reacción de la miokinasa. En fibras musculares rápidas, el AMP se metaboliza a inosina monofosfato (IMP) y amonio (NH3) por la deaminación del AMP. En las fibras lentas, el metabolismo del ADP sigue la dirección hacia la adenosina, vía la reacción 5´-nucleotidasa. El aumento en el AMP y la caída en la CrP, activa la AMP kinasa kinasa,.provocando la fosforilación y activación de la AMP kinasa (AMPK) (Modificado de Hoods, 2001)
Eventos de fosforilaciónLa evidencia es muy basta respecto del efecto positivo del ejercicio en la activación de una variedad de proteínas kinasas que podrían estar potencialmente involucradas en la fosforilación de factores de trascripción.
Los resultados indican que la actividad contráctil per se, más que factores humorales o neurotróficos asociados con el ejercicio, es la que activa la fosforilación de kinasas. Este es un requisito necesario para que la kinasa se vea potencialmente involucrada en la biogénesis mitocondrial inducida por actividad contráctil. Entre las kinasas activadas por la actividad aeróbica se incluyen PKC, ERK-1 y ERK2. MAP kinasas, kinasa p38, AMPK, Raf-1, MAPKAP kinase-2, y proteína kinasa 1 activada por estrés y mitógenos (MSK1), entre otras. Muchos de estos resultados han sido replicados tanto en modelos humanos de actividad contráctil como en roedores, lo cual indica el vasto potencial de la actividad contráctil para alterar las vías de señalización mediadas por kinasas, y subsecuentemente la actividad de unión al ADN de los factores de trascripción, o, alternativamente, la unión al ARN de factores de estabilización mediante modificaciones covalentes.
Una nueva serie de eventos de fosforilación también ocurre a nivel mitocondrial, cuyas implicaciones aún no han sido establecidas. Por muchos años se ha sabido que ciertas subunidades de los complejos piruvato deshigrogenasa y cetoácido de cadena ramificada deshidrogenasa son fosforiladas reversiblemente, lo cual puede cambiar la activación del complejo entero.
Recientemente, se ha encontrado que otras proteínas mitocondriales también se fosforilan. Estas incluyen la subunidad 4 de la proteína COX (citocromo c oxidasa) y las subuinidad de 18 kDa del complejo I. El mecanismo propuesto parece involucrar la presencia de la proteína kinasa A dependiente de AMPc, localizada en la membrana mitocondrial interna. Interesantemente, la fosforilación de la subunidad de 18-kDa activa la respiración mitocondrial mediante un aumento en la actividad del complejo I, mientras que la fosforilación de la subunidad IV reduce la respiración por inhibición de la actividad de la COX. Esta inhibición puede ser revertida por la activación mediada por calcio de una proteína fosfatasa.
Es para remarcar que el calcio no solamente puede activar fosfatasas, sino que también ha sido visto que modula la fosforilación de la subunidad c de la ATPasa F1 F0.
Trascripción de genes nuclearesEn general la trascripción de genes (pasaje de ADN a ARNm) es regulada por proteínas denominadas factores de trascripción, los cuales se unen ciertas regiones de los genes, llamadas regiones regulatorias. Un número de ellos ha sido implicado en la biogénesis mitocondrial, e incluyen NRF-1 y NRF-2 PPAR-a y PPAR-g (peroxisome proliferator-activated receptors-a y -g), Egr-1 y c-Jun, c-Fos, y Sp1. Ha sido demostrado además que la actividad contráctil induce aumentos en el ARNm y/o en los niveles de proteína de muchos de ellos, en concordancia con sus roles en la mediación de cambios fenotípicos en el músculo como resultado del ejercicio.
Se lograría un aumento coordinado en la trascripción de genes mitocondriales y nucleares, si fuese identificado un activador universal de genes. Al respecto, existen sitios para NRF-1 en las regiones promotoras de varios genes nucleares, incluyendo el factor de trascripción mitocondrial A (Tfam), como también de ALAs, la enzima limitante de la síntesis del grupo hemo. Sin embargo los sitios para NRF-1 no se han encontrado universalmente, por ej. no todas de las 10 subunidades de la COX codificadas por el núcleo, poseen sitios para NRF-1.
Recientemente se ha descubierto un coactivador de PPAR-g llamado PGC-1 que podría ser un potencial candidato regulatorio. La sobreexpresión de PGC-1 ha demostrado inducir índices morfológicamente mensurables del contenido mitocondrial, como también la expresión del ARNm de un número de genes nucleares en las células musculares, incluyendo el NRF-1, NRF-2a, Tfam, UCP-2, subunidades de COX codificadas nuclear y mitocondrialmente, y el citocromo c. El incremento de la expresión de Tfam, conduce a un aumento del ADNmt. Este efecto se debe probablemente al resultado de la coactivación del NRF-1 ejercida por PGC-1, sobre el promotor de Tfam. Evidencia anexa provista por otros experimentos, sugieren que el NRF-1 es necesario para el efecto de PGC-1 sobre la biogénesis mitocondrial, y además otro trabajo demostró que el PGC-1 fue capaz de coactivar una segunda proteína, la PPAR-a, un receptor conocido por regular la expresión de enzimas involucradas en la oxidación de ácidos grasos como la acetil-CoA de cadena media.
La biogénesis mitocondrial requiere de la cooperación del genoma nuclear y mitocondrialUno de los aspectos más interesantes de la síntesis mitocondrial es que requiere la acción coordinada de los genes nucleares y de los mitocondriales.
La mitocondria posee numerosas copias de una molécula pequeña de ADN circular (ADNmt) que comprende aproximadamente 16.659 nucleótidos. A partir de ese ADN, puede sintetizar 13 proteínas que forman parte esencial de la cadena de transporte de electrones, sin embargo la gran mayoría de las proteínas mitocondriales provienen de la información codificada en el ADN del núcleo celular.
Los mecanismos de señalización, como los que son activados por el ejercicio, inician la activación de factores de trascripción, que aumentan la producción de ARNm a partir del DNA nuclear y mitocondrial. Los productos de los genes nucleares son traducidos en el citosol como "proteínas precursoras" con una secuencia señal (targeting) localizada en el extremo amino terminal o en una secuencia interna proteínas precursoras. Estas proteínas precursoras interaccionan con otras proteínas llamadas chaperonas (ej: proteína de shock térmico de 70-kDa o HSP70 y el factor mitocondrial de estimulación de la importación o MSF), que mantienen la proteína recién sintetizada en forma desplegada y la dirigen a la maquinaria de importación de la membrana mitocondrial externa, donde hay un grupo de proteínas conocidas como complejo Tom. El precursor se transfiere luego a través de la membrana externa, por el espacio intermembranoso, hacia otras proteínas llamadas traslocasas, o complejo Tim, de la membrana mitocondrial interna.
Ciertos componentes intramitocondriales, empujan al precursor hacia la matriz mitocondrial, clivan la secuencia de reconocimiento de destino, y la proteína termina replegándose en la conformación madura. Situaciones de estrés fisiológico como la actividad contráctil y la hormona tiroidea aceleran la importación de proteína hacia la mitocondria, coincidentemente con el aumento en la expresión de algunos componentes de la maquinaria de importación. En forma contraria, deterioros en el proceso de importación puede causar disfunción mitocondrial y enfermedad.
Ejercicio y plasticidad mitocondrial¿Qué señales disparan la biogénesis mitocondrial?
Está bien establecido que el ejercicio de resistencia, empleando una duración apropiada por día, cierta frecuencia semanal y una intensidad submáxima por sesión de entrenamiento, puede producir un aumento del contenido mitocondrial, usualmente en un rango de 50 al 100% en 6 semanas. Esto produce una mejora en la performance de resistencia, mayormente en forma independiente de los cambios mucho más pequeños inducidos por el ejercicio en el consumo máximo de oxígeno. La biogénesis mitocondrial puede ser replicada utilizando modelos artificiales de ejercicio, como la actividad contráctil crónica producida por estimulación eléctrica del nervio motor. También puede ser el resultado de otras condiciones fisiológicas, incluyendo el tratamiento con hormona tiroidea, actuando a través de receptores nucleares y mitocondriales. La respuesta del músculo a la hormona tiroidea es específica según el tipo de fibra, dependiendo en parte de las diferencias en la distribución de los receptores nucleares para esta hormona.
Las vías que utiliza el tratamiento con hormona tiroidea y las que utiliza la actividad contráctil para producir biogénesis mitocondrial son independientes entre sí, y esto ha sido evidente porque aumentos en el contenido mitocondrial son aún notables en los músculos de animales que han sido tiroidectomizados y sometidos a actividad contráctil crónica. De cualquier manera, ambas condiciones parecen producir un incremento en la masa mitocondrial en las células musculares, resultando en una capacidad mayor para el consumo de oxígeno por gramo de tejido.
Mediciones de proteínas marcadoras como la actividad del citocromo c o de la citocromo c oxidasa (COX), han permitido a los fisiólogos del ejercicio reconocer que, en respuesta a un estímulo de ejercicio constante, se requieren 6 semanas de entrenamiento de la resistencia (endurance) para alcanzar un contenido mitocondrial estacionario mayor. Este tiempo depende del tipo de fibra que está siendo reclutado, como también de la duración, frecuencia e intensidad de la sesión de ejercicio.
Las adaptaciones mitocondriales no ocurrirán en las células musculares esqueléticas que no hayan sido reclutadas durante la sesión de ejercicio, consistentemente con la idea de que el estímulo para la biogénesis se origina en el músculo en contracción, independientemente de otras influencias. Un corolario de esto es que los programas de entrenamiento de la resistencia deben ser apuntados, en parte, hacia el reclutamiento de unidades motoras conteniendo fibras tipo IIb (blancas rápidas). Interesantemente, el entrenamiento en contra de resistencia, el cual sí recluta unidades motoras fatigables rápidamente, no conduce a una adaptación mitocondrial. Como la muy alta intensidad y la corta duración de la mayoría de los programas de entrenamiento contra resistencia representan un estímulo fuerte para la síntesis de proteínas miofibrilares, conduciendo a la hipertrofia, el contenido mitocondrial en las fibras musculares agrandadas, podría estar "diluido". Esto incrementa la distancia de difusión del oxígeno y de los sustratos, y no representa una adaptación favorable con respecto a la performance en el entrenamiento de endurance.
El período de tiempo aproximado de 6 semanas requerido para lograr una nuevo estado estacionario (steady-state) en el contenido mitocondrial en respuesta al entrenamiento de resistencia, claramente no refleja los eventos moleculares tempranos que en última instancia conduce a cambios morfológicamente mensurables.
Las proteínas mitocondriales se recambian con una vida media de 1 semana luego del inicio de un nuevo nivel de actividad contráctil muscular. Esto significa que se requiere un estímulo de ejercicio continuo para mantener el contenido de mitocondrias en un nivel elevado luego de un período de entrenamiento; de otra manera, se producirá la pérdida de la capacidad oxidativa y de resistencia.
Es interesante hacer notar que los cambios en el contenido mitocondrial de fosfolípidos, ocurren con una vida media aún más corta (4 días), sugiriendo que el ensamblado y/o la degradación de la organela podría ser iniciada por cambios en la composición de fosfolípidos.
Por ejemplo, en el músculo con deficiencia de hierro, se alteran las velocidades normales de síntesis de citocromo (ya que se necesitan grupo hemo) y su incorporación a la membrana interna, sin embargo cuando se le impone una actividad contráctil a ese músculo, la misma parece conducir a la síntesis continua de lípidos de membrana, produciendo un volumen mitocondrial aumentado, con un contenido proteico notablemente reducido y anormal. Estos datos sugieren que la síntesis de proteínas y de fosfolípidos no necesariamente están acopladas durante la biogénesis mitocondrial inducida por actividad contráctil. Los datos también sugieren que la proporción de lípidos a proteínas en la mitocondria debe cambiar un poco durante la transición tanto hacia contenidos mitocondriales más altos o más bajos. Este cambio parece ser sutil como resultado del entrenamiento o de la estimulación crónica, y en la mayoría de los casos parece no afectar la velocidad de consumo de oxígeno por unidad de volumen mitocondrial. Sin embargo, podría ser mucho más evidente bajo condiciones de steady-state en tejidos que posean contenidos mitocondriales ampliamente divergentes, como el corazón y el músculo esquelético blanco.
Cambios en el metabolismo celular producidos por la biogénesis mitocondrial durante el ejercicio de resistenciaLa notable mejora en la performance de la resistencia que resulta de la biogénesis mitocondrial es una consecuencia de cambios en el metabolismo muscular durante el ejercicio. Durante la contracción muscular intensa, la concentración de ADP libre (ADPl) aumenta. Este incremento conduce el equilibrio de la reacción en la que interviene la creatina fosfokinasa (CPK) hacia la formación de ATP y creatina (ver fig 1). El ADPl es también un sustrato y efector alostérico en la vía glucolítica, y controla la respiración mitocondrial activa. Como el entrenamiento de la resistencia incrementa el contenido mitocondrial del músculo esquelético sin grandes efectos sobre la CPK o las enzimas glucolíticas, parece razonable asumir que una fracción mayor del requerimiento de energía para un determinado trabajo de esfuerzo, derivará ahora del metabolismo aeróbico.
Esto ha sido referido como una sensibilidad mayor de la respiración mitocondrial al ADPl, ya que se requiere un incremento menor en la concentración de este metabolito, para lograr el mismo nivel de consumo de oxígeno. Este incremento reducido en el ADPl atenuará la glucólisis y la formación de ácido láctico, y se producirá una economía de fosfocreatina. Además, la formación de ADP ocurrirá a una velocidad menor. En el músculo rápido, con una alta actividad de la AMP-deaminasa, la síntesis de amoníaco y de IMP se reducirá. En el músculo lento, que posee menos AMP-deaminasa y actividades mayores de 5´-nucleotidasas, la menor formación de AMP será traducida en una conversión reducida a adenosina (ver fig 1). Con la declinación más pequeña de creatinafosfato, una síntesis reducida de AMP, conducirá a una activación menor de la AMP kinasa (AMPK), un factor que podría ser importante para la reducción inducida por el entrenamiento de la traslocación de GLUT4 (transportador de glucosa) hacia la membrana plasmática. Esto, en coincidencia con actividades aumentadas de enzimas mitocondriales de la b-oxidación, predispone al individuo hacia una oxidación lipídica mayor durante el ejercicio.
Un estudio investigó la importancia de la creatina kinasa (CK) en la regulación de la respiración mitocondrial muscular, en humanos, dependiendo de su nivel de actividad física. Los voluntarios fueron clasificados en sedentarios, activos o atléticos, de acuerdo al índice de actividad total determinado por el cuestionario de Baecke, en combinación con valores máximos de oxígeno (VO2 máximo), expresado en ml min(-1) kg(-1)). Todos los voluntarios realizaron test de ejercicio incremental en cicloergómetro a su VO2 máx y VO2 en el umbral ventilatorio (VT).
Se tomaron muestras de biopsias musculares del vastus lateralis y la respiración mitocondrial fue evaluada en una celda oxigráfica, en ausencia (V(0)) o en presencia (V(máx)) de concentración saturada de ADP. Mientras que V(0) fue similar, V(máx.) difirió entre los grupos. V(máx.) se correlacionó con el VO2 máx y con VT. Hubo un grado significativamente mayor de acoplamiento entre la oxidación y la fosforilación (V(máx.)/V(0)) en los individuos atletas. La Km (constante de equilibrio de una reacción) mitocondrial para el ADP fue significativamente mayor en los sujetos atléticos. La activación de la CK mitocondrial por la adición de creatina, indujo un marcado descenso en la Km (a menor Km, mayor es la afinidad por el sustrato, es decir que la reacción se ve facilitada hacia la formación del producto) en los individuos atléticos solamente, indicación de un eficiente acoplamiento de la CK mitocondrial a la refosforilación del ADP en dichos sujetos.
Se ha sugerido que la performance aeróbica aumentada requiere una mejora tanto de la capacidad muscular oxidativa como de los mecanismos de control respiratorio, dando muestra de la importancia de la coordinación temporal de los flujos de energía por la CK, para más eficacia.
Otros estudios también muestran que después del entrenamiento, la sensibilidad de la mitocondria al ADP, disminuye, mientras que el efecto de la creatina en la respiración se encuentra aumentado, lo cual resulta en una mejora en el control de la producción de energía aeróbica.
El incremento del contenido mitocondrial producido por el ejercicio de resistencia no solamente conduce a una producción menor de lactato, sino también a una mejora en lavado del mismo.
El transporte de lactato hacia la mitocondria puede ocurrir vía el transportador mitocondrial monocarboxilado (MCT1), por lo tanto el aumento del contenido de mitocondrias aumentará la oxidación del lactato, produciendo velocidades menores de liberación desde el músculo. Además, cuanto mayor es el contenido de mitocondrias por gramo de músculo, menor es la velocidad de respiración requerida por mitocondria para cualquier carga de trabajo dada. Esto predice una reducción en los niveles de las especies reactivas de oxígeno (ROS), potencialmente peligrosas, aún a una velocidad constante de consumo de oxígeno por gramo de músculo. La combinación de la producción reducida de ROS, con un aumento de las enzimas protectoras, sustenta la idea de un beneficio adicional de la actividad contráctil crónica: además de la capacidad oxidativa aumentada, también atenúa el daño potencial que podrían producir las ROS en las proteínas, los lípidos y el ADN.
Contrariamente, un contenido reducido de mitocondrias, como es evidente en algunos tejidos, como consecuencia del envejecimiento, se correlaciona con una mayor tendencia al daño inducido por ROS, particularmente sobre el ADNmt. Este hecho pone de manifiesto una razón potencialmente importante para mantener el contenido mitocondrial del músculo con un programa de actividad física regular durante el proceso de envejecimiento.
ConclusionesEl ejercicio (es decir, la actividad contráctil) produce cambios fisiológicos, bioquímicos y moleculares dependientes del tiempo. La magnitud de dichas modificaciones es dependiente de la intensidad y duración del estímulo del ejercicio aeróbico. De esta forma es posible alterar el fenotipo muscular, de una fibra blanca con baja capacidad oxidativa a una roja altamente oxidativa. Este cambio conduce a una adaptación metabólica que atenúa algunas señales de estrés impuestas por una sesión intensa de ejercicio (por ejemplo la producción y remoción de lactato), actuando en cierto modo como un feedback negativo. Por lo tanto las señales para producir una futura biogénesis mitocondrial se presume que se ven reducidas en magnitud una vez que la adaptación ha tenido lugar.
El entrenamiento de la resistencia (endurance) conduce a incrementos del volumen mitocondrial (volumen de la célula ocupado por mitocondrias) de hasta el 100% con entrenamiento de unas pocas semanas en sujetos no entrenados. El entrenamiento de la fuerza tiene un impacto mayor en el volumen de las miofibrillas musculares, sin embargo la mitocondria aparece relativamente sin cambios.
El interés científico actual por la función mitocondrial, su biogénesis y la expresión genética en células musculares no ha sido nunca tan intenso. Esto se debe a que la mitocondria provee un excelente modelo adaptativo para el estudio de la biogénesis de organelas en las células de mamífero y porque se sabe ahora, que está implicada en una amplia variedad de enfermedades.
Referencias
Adhihetty PJ et al. Plasticity of skeletal muscle mitochondria in response to contractile activity. Exp Physiol 2003;88(Pt 1):99-107.
Dubouchaud H et al. Endurance training, expression, and physiology of LDH, MCT1, and MCT4 in human skeletal muscle. Am J Physiol Endocrinol Metab 278: E571-E579, 2000.
Dudley GA et al. Influence of exercise intensity and duration on biochemical adaptations in skeletal muscle. J Appl Physiol 53: 844-850, 1982.
Dudley GA et al. Influence of mitochondrial content on the sensitivity of respiratory control. J Biol Chem 262: 9109-9114, 1987.
Essig DA. Contractile activity-induced mitochondrial biogenesis in skeletal muscle. Exerc Sport Sci Rev 24: 289-319, 1996.
Enriquez JA et al. Direct regulation of mitochondrial RNA synthesis by thyroid hormone. Mol Cell Biol 19: 657-670, 1999.
Freyssenet D et al. Calcium-dependent regulation of cytochrome cgene expression in skeletal muscle cells: identification of a protein kinase c-dependent pathway. J Biol Chem 274: 9305-9311, 1999.
Gordon JW et al. Plasticity in skeletal, cardiac, and smooth muscle selected contribution: effects of contractile activity on mitochondrial transcription factor A expression in skeletal muscle. J Appl Physiol 90: 389-396, 2001.
Gore M et al. Endurance training alters antioxidant enzyme gene expression in rat skeletal muscle. Can J Physiol Pharmacol 76: 1139-1145, 1998.
Gottlieb RA. Mitochondria: execution central. FEBS Lett 482: 6-12, 2000.
Grey JY et al. Tom20-mediated mitochondrial protein import in muscle cells during differentiation. Am J Physiol Cell Physiol 279: C1393-C1400, 2000.
Herzig RP et al. Sequential serum-dependent activation of CREB and NRF-1 leads to enhanced mitochondrial respiration through the induction of cytochrome c. J Biol Chem 275: 13134-13141, 2000.
Holloszy JO and Coyle EF. Adaptations of skeletal muscle to endurance exercise and their metabolic consequences. J Appl Physiol 56: 831-838, 1984.
Holloszy JO and Winder WW. Induction of d-aminolevulinic acid synthetase in muscle by exercise or thyroxine. Am J Physiol Regulatory Integrative Comp Physiol 236: R180-R183, 1979.
Hood DA et al. Mitochondrial adaptations to chronic muscle use: effect of iron deficiency. Comp Biochem Physiol 101A: 597-605, 1992.
Hood DA at al. Effect of thyroid status on the expression of metabolic enzymes during chronic stimulation. Am J Physiol Cell Physiol 263: C788- C793, 1992.
Hood DA et al. Assembly of the cellular powerhouse: current issues in muscle mitochondrial biogenesis. Exerc Sport Sci Rev 28: 68-73, 2000.
Hood DA. Plasticity in Skeletal, Cardiac, and Smooth Muscle. Invited Review: Contractile activity-induced mitochondrial biogenesis in skeletal muscle. J Appl Physiol 90: 1137-1157, 2001.
Hood DA et al. Mitochondrial biogenesis and the role of the protein import pathway. Med Sci Sports Exerc 2003;35(1):86-94
Hoppeler H. Exercise-induced ultrastructural changes in skeletal muscle. Int J Sports Med 7: 187-204, 1986.
Hoppeler H et al. Endurance training in humans: aerobic capacity and structure of skeletal muscle. J Appl Physiol 59: 320-327, 1985.
Hoppeler H, Fluck M. Plasticity of skeletal muscle mitochondria: structure and function. Med Sci Sports Exerc 2003;35(1):95-104.
Jones TE et al. Exercise induces an increase in muscle UCP3 as a component of the increase in mitochondrial biogenesis. Am J Physiol Endocrinol Metab 2003;284(1):E96-101.
Koehler CM. Protein translocation pathways of the mitochondrion. FEBS Lett 476: 27-31, 2000.
Kraniou Y et al. Effects of exercise on GLUT-4 and glycogenin gene expression in human skeletal muscle. J Appl Physiol 88: 794-796, 2000.
Kubis H-P et al. Adult fast myosin pattern and Ca 2+-induced slow myosin pattern in primary skeletal muscle culture. Proc Natl Acad Sci USA 94: 4205-4210, 1997.
Mitsui T et al.Chronic corticosteroid administration causes mitochondrial dysfunction in skeletal muscle. J Neurol 2002;249(8):1004-9.
Schrauwen P et al.Etomoxir-induced increase in UCP3 supports a role of uncoupling protein 3 as a mitochondrial fatty acid anion exporter. FASEB J 2002;16(12):1688-90.
Tonkonogi M, Sahlin K.Physical exercise and mitochondrial function in human skeletal muscle. Exerc Sport Sci Rev 2002;30(3):129-37.
Wu H et al. Regulation of mitochondrial biogenesis in skeletal muscle by CaMK. Science 2002; 296(5566):349-52.
Zoll J et al. Physical activity changes the regulation of mitochondrial respiration in human skeletal muscle. J Physiol 2002;543(Pt 1):191-200.
| |
|
|---|---|
|
revista
digital · Año 9 · N° 61 | Buenos Aires, Junio 2003 |
|