
|
|||
|---|---|---|---|
|
|
Marcadores fisiológicos de overtraining (parte 1): análise de um protocolo para ratos ‘wistar’submetidos a treinamento de corrida contínua Marcadores fisiológicos de sobreentrenamiento (parte 1): análisis de un protocolo para ratas ‘wistar’ sometidas a entrenamiento de carrera continua Physiological markers of overtraining (part 1): analysis of a protocol for rats ‘wistar’ submitted to continuous exhaustive training |
|
|
|
*Doutor em Biologia Funcional e Molecular. Docente do curso de Fisioterapia do Instituto de Ensino Superior de Itapira, IESI **Mestranda em Ciências Médicas (UNICAMP). Coordenadora do curso de graduação em Fisioterapia do Instituto de Ensino Superior de Itapira, IESI ***Graduado em Educação Física, Faculdades Integradas Metropolitanas de Campinas, METROCAMP ****Professora Associada do Instituto de Biologia. Departamento de Bioquímica, Universidade Estadual de Campinas, UNICAMP |
Joaquim Maria Ferreira Antunes Neto* Bruna Bergo Nader** Rodrigo José Battibugli Rivera*** Caio Cesar Donadon*** Denise Vaz de Macedo**** (Brasil) |
|
|
|
Resumo O objetivo deste trabalho foi comparar níveis de alterações fisiológicas em ratos Wistar submetidos a um protocolo de overtraining por meio de corrida contínua em esteira rolante automatizada. Os animais passaram por três fases de treinamento, que variaram da adaptação, manutenção e incremento de sobrecarga. Ao final do procedimento, os animais chegaram a correr quatro vezes por dia, cinco vezes por semana. Os parâmetros estudados foram concentrações plasmáticas de ácido úrico, creatinina e uréia (mg/dL). Todos apresentaram aumentos significativos em algum período de incremento de sobrecarga (p<0.05), sugerindo que o acompanhamento destes marcadores pode contribuir para a detecção ou estabelecimento de indícios de instalação de uma situação de overtraining. Unitermos: Overtraining. Ácido úrico. Creatinina. Uréia. Corrida contínua. Experimentação animal.
Abstract The objective of this study was to compare levels of physiological changes in rats subjected to an overtraining through continuous running on a treadmill automated. The animals underwent three phases of training, ranging from adjustment, maintenance and increased overload. At the end of procedure, the animals reached run four times a day, five times per week. The parameters studied were plasma concentrations of uric acid, creatinine and urea (mg/dL). All data showed significant increases in some period of increased overload (p <0.05), suggesting that monitoring of these markers may contribute to the establishment of detection or indication of installing a state of overtraining. Keywords: Overtraining. Uric acid. Creatinine. Urea. Continuous running. Animal experimentation.
|
|||
|
|
EFDeportes.com, Revista Digital. Buenos Aires - Año 17 - Nº 170 - Julio de 2012. http://www.efdeportes.com/ |
|
|
1 / 1
Introdução
A falta de sincronismo entre volume de exercícios, intensidade de esforço e recuperação, estabelecida por uma metodologia que visa o condicionamento e elevação das capacidades físicas de um atleta, é fator determinante para indução de sobrecarga dos sistemas fisiológicos, o que tente a ocasionar uma superatividade dos mecanismos metabólicos e funcionais do organismo e até mesmo, em situação extrema, a desintegração dos sistemas de defesa deste (Bompa, 1990). Tais fatores são elementos relevantes na instalação da síndrome de overtraining. Os sinais associados a esta síndrome são alta fadigabilidade e outros distúrbios, tais como respostas inflamatórias, aumento de citocinas circulantes no sangue, desbalanço nutricional, distúrbios hormonais, indisposição ao treinamento físico, que resultam em diminuição do rendimento e aumento na susceptibilidade a lesões (Johnson, Thiese, 1992; Kibler et al., 1992; Lehmann et al., 1993; Pascoe et al., 1993; Eichner, 1995; Hooper et al., 1995; Kuipers, 1996; Traverso et al., 1996; Tenenbaum, 1996; Tiidus,1998; Lehmann et al., 1998). O overtraining, uma vez instalado, não é facilmente revertido, podendo levar várias semanas ou meses para ocorrer a recuperação (Kuipers, Keizer, 1988).
Dentro do contexto neuroendócrino de classificação da síndrome do overtraining, Lehmann e colaboradores (1998) relataram a possibilidade de haver duas formas da síndrome: tipo parassimpática (aumento do tônus parassimpático em repouso e no exercício), geralmente uma conseqüência do desbalanço entre alto volume de treinamento de longa duração e pequeno tempo para regeneração (esportes de endurance); e tipo simpática (aumento do tônus simpático em repouso), quando há, sobretudo, sessões de treinamentos mais intensivas e intermitentes. Embora os mecanismos fisiopatológicos de ambos os tipos ainda não sejam totalmente conhecidos, surgiu a hipótese de que eles refletiriam estágios diferentes da própria síndrome; durante o estágio inicial, o sistema simpático seria continuamente “alertado”, enquanto que durante os estágios mais avançados, a atividade do sistema simpático seria inibida, resultando em uma prevalência do sistema parassimpático (Kuipers, 1996).
Dosagens plasmáticas de ácido úrico, creatinina e uréia são muito utilizadas em experimentações que correlacionam o esforço físico a alterações catabólicas no organismo (Lapin et al., 2007; Córdova, Navas, 2000). Porém, poucos estudos desenvolvem procedimentos metodológicos para intervenção em situação de overtraining, sobretudo pela dificuldade de relacionar as multivariáveis fisiológicas instaladas a uma condição tão complexa de delineamento (Antunes Neto et al., 2008 a, b). A tentativa de compreender melhor a modulação de parâmetros fisiológicos por intermédio de experimentação animal é valida, pois há a possibilidade de elaboração de protocolos que não seriam possíveis de controle através da participação de seres humanos. Assim, visamos contribuir no aprofundamento sobre determinação de indicadores satisfatórios de diagnóstico do quadro de overtraining.
Materiais e métodos
-
Animais. Utilizamos ratos machos Wistar, obtidos do Centro de Bioterismo da UNICAMP, com 2 meses de idade no início dos protocolos de exercícios. Durante toda a execução dos protocolos, os ratos foram mantidos em ambiente controlado, com temperatura de 18 - 22°C e fotoperíodo invertido de 12 horas, estando providos de ração de laboratório e água ad libitum.
Proposta de Protocolo Contínuo de Corrida para Indução de Overtraining
Os ratos foram submetidos a sessões de corrida em esteira rolante automatizada com inclinação de -3°. Porém, antes da realização do respectivo protocolo, realizamos um período de adaptação (1 semana) do animal com o equipamento, onde eles chegaram a correr por 5 minutos numa velocidade de 10m/min. Esta semana inicial teve como finalidade separar os animais que corriam voluntariamente daqueles que se recusavam a correr. O protocolo de exercício contínuo teve onze semanas de duração, dividido da seguinte forma:
-
primeiras quatro semanas (adaptação): aumento na carga de esforço;
-
4a. a 8a. semana (manutenção): exercícios diários com a mesma carga de esforço alcançada ao final da 4a semana;
-
semana 2x/dia (9ª. Semana – incremento 1): uma semana de exercícios com a mesma carga das semanas anteriores, 2 vezes ao dia, com intervalo de 4 horas entre eles.
-
semana 3x/dia (10a. semana – incremento 2): uma semana com 3 sessões de treino ao dia, com intervalo de 3 horas entre eles;
-
semana 4x/dia (11a. semana – incremento 3): uma semana com 4 sessões de treino ao dia, com intervalo de 2 horas entre eles.
Tabela 1. Protocolo de treinamento contínuo para indução de overtraining.

-
Preparo da amostra. O plasma foi obtido após centrifugação, por 10 min a 3000 rpm, do sangue restante. O plasma foi separado e armazenado a -80°C após tratado com butilhidroxitolueno (BHT) 90mM, este que atua como um seqüestrador específico de espécies reativas de oxigênio.
-
Dosagens de parâmetros metabólicos.
As
análises de creatinina, uréia e ácido úrico foram realizadas no aparelho
Autolab PM 4000, através de kits de análises clínicas específicos: TGO –
kit QUIMIAST, uréia e creatinina – kit QUIMIURE, ambos da Böehringer
Mannheim, e ácido úrico – kit específico Roche.
-
Apresentação gráfica e análises estatísticas.
Os
gráficos estão apresentados no modelo box plot (software S-Plus). O box
plot ordena os dados pela mediana e os divide em quartis. Ou seja, acima e
abaixo da mediana encontram-se 50% dos dados obtidos, 25% acima da mediana e
25% abaixo dela e as duas linhas tracejadas representam os 50% dos valores
mais distantes. Os dados que se diferenciam extremamente do comportamento
geral são colocados para fora dos quartis e são chamados de outliers
ou valores extremos. Optamos por apresentar os resultados obtidos na forma de box
plot por entendermos que esse tipo de gráfico descreve melhor o
comportamento do grupo de animais frente às várias situações de
exercício, permitindo uma melhor visualização da variabilidade dos dados. O
interessante desta forma de apresentação gráfica é a possibilidade de
poder visualiar a disposição de todo o grupo, conseguindo, assim, situar a
maior concentração dos resultados (Mcgill et al., 1978). As análises
estatísticas foram feitas através do programa GraphPad Instat® (San Diego,
CA). Utilizamos o teste ANOVA para amostras pareadas e, como pós-teste, foi
adotado o teste de Tukey. Valores de p<0.05 foram considerados
significativos. Vale ressaltar que a apresentação gráfica ressalta valores
da mediana, enquanto que o tratamento estatístico considera média e
desvio-padrão.
Resultados
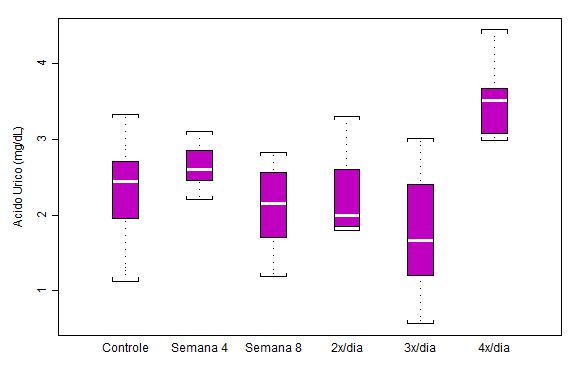
Figura 1. Concentração plasmática de ácido úrico (mg/dL) em ratos submetidos a protocolo de treinamento
contínuo por 8 semanas e ao aumento na freqüência diária de corrida (n = 8 para cada semana)
Os dados da concentração plasmática de ácido úrico revelam que esse marcador começa a mostrar alteração mais acentuada somente na última etapa de treinamento. Nota-se, ainda, que o último box (Semana 4 x/dia) torna-se mais compacto, o que significa homogeneidade do resultado, com um aumento considerável chegando a 65.1% em relação ao valor do grupo Semana 8 (p<0.001). Demais alterações estatísticas significativas: p<0.01 4x/dia em relação ao grupo controle; p<0.01 4x/dia em relação a 2x/dia; p<0.001 4x/dia em relação a 3x/dia.
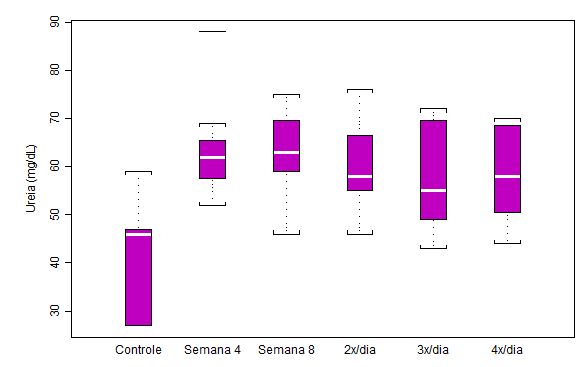
Figura 2. Concentração plasmática de ácido úrico (mg/dL) em ratos submetidos a protocolo de treinamento
contínuo por 8 semanas e ao aumento na freqüência diária de corrida (n = 8 para cada semana)
Nota-se que a concentração de uréia no plasma aumentou com o início do treinamento (análise Semana 4), indicando um aumento na taxa catabólica protéica. Esse aumento na concentração de uréia chegou a 34.7%. Os valores medianos de uréia não sofreram alterações drásticas a partir da análise da Semana 4, e a variação na distribuição dos resultados em cada momento de análise, apesar de bastante heterogênea, manteve-se similar até a análise 4x/dia. Houve aumento significativo de p<0.05 dos demais grupos em relação ao controle.
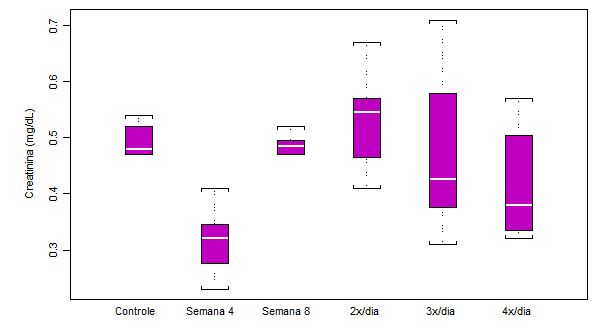
Figura 3. Concentração plasmática de creatinina (mg/dL) em ratos submetidos a protocolo de treinamento
contínuo por 8 semanas e ao aumento na freqüência diária de corrida (n = 8 para cada semana)
Perceba que concentração de creatinina (mg/dL) diminuiu com o início do treinamento (análise Semana 4), indicando adaptação. Esse decréscimo na concentração de creatinina chegou a 33.4% em relação ao controle. Houve uma tendência de retorno na semana 8 para valores próximos do grupo controle, porém mostrando grande heterogeneidade nos resultados a partir da análise 2x/dia. Os valores de significância estatística foram: p<0.001 controle em relação a semana 4; p<0.01 Semana 4 em relação a Semana 8; p<0.01 semana 4 em relação a 3x/dia.
Discussão
Para melhor compreensão da temática, buscaremos descrever as bases fisiológicas de formação do ácido úrico, creatinina e uréia. Posteriormente, analisaremos os resultados obtidos e as possíveis considerações decorrentes da aplicação do protocolo descrito.
Os nucleotídeos participam de importantes etapas do metabolismo, tais como fornecimento de energia, funcionam como agentes de fosforilações (ATP), mediadores fisiológicos (AMPc), componentes de co-enzimas (NAD+, NADP+, FAD, CoA) e intermediários ativados (UDP-glicose, UDP-galactose). Além disso, sua degradação produz precursores de vitaminas e alcalóides (derivados de xantina). A hidrólise dos ácidos nucléicos, que ocorre no duodeno, é realizada por enzimas específicas chamadas DNases e RNases, e resulta em mononucleotídeos. Estes, por sua vez, sofrem a ação de fosfatases, que os quebram em bPO4- e nucleosídeos (um açúcar ligado a uma base nitrogenada). Finalmente, os nucleosídeos são quebrados por nucleosidases, resultando em bases púricas e pirimídicas e açúcares D-ribose e D-desoxirribose. Os nucleotídeos derivados da hidrólise dos ácidos nucléicos são catabolisados, formando açúcares, fosfato e bases púricas e pirimídicas. No homem e nos primatas, as bases púricas são catabolisadas a ácido úrico (Figura 4). As bases pirimídicas são catabolisadas a CO2 e NH3.
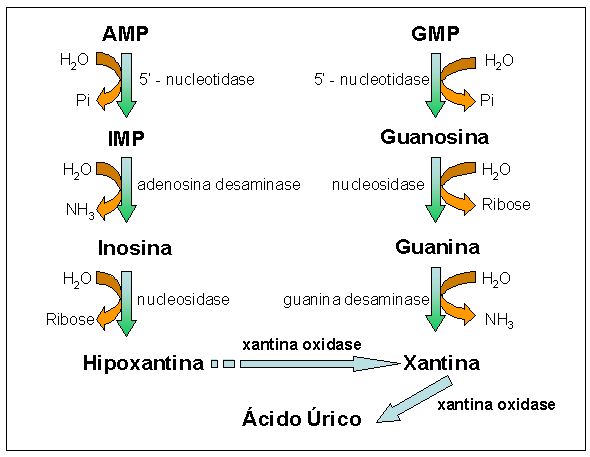
Figura 4. Processo de catabolização de nucleotídeos púricos a ácido úrico
A creatinina é originária da glicociamina (guanidoacetato) produzida largamente nos rins, intestino delgado, pâncreas e, principalmente, no fígado (Figura 5). No fígado, esta glicociamina é convertida em creatina, que é fosforilada no músculo em fosfocreatina, pela ação da enzima creatina quinase (CK). A fosfocreatina corresponde à reserva de fosfato que o músculo recorre para a síntese de ATP em anaerobiose, juntamente com a glicólise, liberando, então, o anidrido da creatina que corresponde à creatinina.
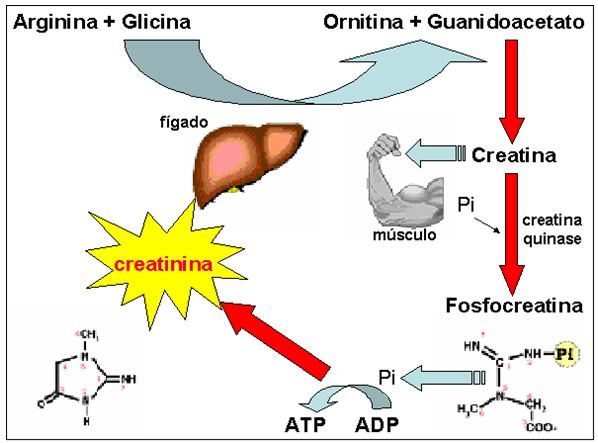
Figura 5. Síntese da creatinina. No fígado, a arginina é adicionada a glicina gerando ornitina e guanioacetato que, por sua vez, é metilado (ainda no fígado)
gerando a creatina. Nos músculos, a creatina é adicionada de fosfato gerando a fosfocreatina, que é uma reserva de fosfato para o processo
metabólico muscular. O consumo do fosfato da fosfocreatina libera a creatinina (que também pode ser formada por hidrólise direta da creatina)
São produzidos, diariamente, cerca de 1,5 g de creatinina em resposta à atividade muscular, podendo atingir até 2,0 g/dia em homens com 70 kg de peso e um mínimo de 900mg/dia em mulheres com 60 kg de peso, em virtude de a formação de creatinina estar relacionada com a massa corpórea de cada indivíduo. A creatinina produzida no metabolismo muscular não é reabsorvida nos túbulos, sendo que em virtude de seu baixo limiar de excreção, as concentrações plasmáticas permanecem em níveis muito baixos de cerca de 0,5 a 1,5 mg/dL.
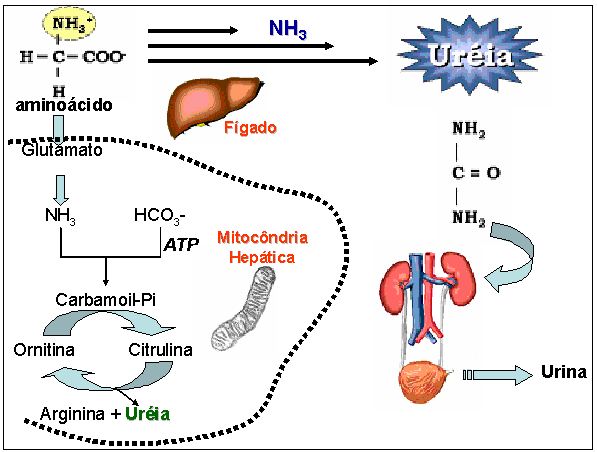
Figura 6. Representação esquemática simplificada do ciclo da uréia. Ciclo hepático da síntese da uréia. O CO2 é adicionado à primeira
molécula de NH3 e ao aminoácido não codificado ornitina na mitocôndria, gerando o aminoácido citrulina que se liga com uma segunda molécula
de NH3, no citoplasma, originando o aminoácido arginina que é quebrado em uréia e ornitina, que penetra na mitocôndria e reinicia o ciclo
Diariamente, são produzidos em torno de 50 a 100g de uréia a partir do metabolismo do grupamento amino dos aminoácidos que ocorre no fígado (Figura 6), como uma forma atóxica de transportar amônia (esta, extremamente tóxica). A uréia é prontamente excretada pelos rins, havendo reabsorção de parte da uréia filtrada, mantendo-se a sua concentração plasmática (uremia) em torno de 10 a 40 mg/dL. Qualquer diminuição na função excretora do rim revela um aumento da uremia (hiperuremia) e a conseqüente diminuição da uréia urinária. Porém há possibilidade de uma uremia de mais de 50 mg/dl (hiperuremia) não associada a causas patológicas renais, mas sim por uma alimentação hiperprotéica antecedendo o teste laboratorial, o que fará a uremia aumentar sem, porém, haver a diminuição da uréia urinária. A hiperuremia pode estar patologicamente relacionada a outras causas não renais, como no choque traumático ou hemorrágico, desidratação aguda, descompensação cardíaca, infecção aguda, toxemia, queimaduras extensas, febre. Também são observados aumento na uremia decorrentes de obstrução pós-renal no nível da uretra (cálculos, coágulos, tumores pélvicos), vesical (tumores compressivos, cálculos) e até mesmo na uretra. Hiperuremia devido a causas exclusivamente renais são observadas na glomerunefrite (podendo atingir taxas de até 300 mg/dL), esclerose renal, tuberculose renal, rim policístico, nefropatias, saturnismo.
A formação dos compostos analisados em nosso estudo relaciona-se com eventos catabólicos de bases púricas e aminoácidos. A elevação, portanto, dos metabólitos (ácido úrico, creatinina e uréia) indica que a intensidade do esforço empregada neste modelo de treinamento contínuo solicita demanda adicional de substratos energéticos, sobretudo de aminoácidos, os quais seguirão para rotas metabólicas (gliconeogênese) relevantes para a continuidade do esforço e dos processos de recuperação.
No caso das análises de concentração de ácido úrico no plasma, observamos que não houve uma alteração drástica dos valores de mediana até o momento 3x/dia. Porém, obtivemos uma elevação considerável em concentração plasmática de ácido úrico na análise 4x/dia. Este fato sugere que a partir da realização de 4 sessões diárias de corrida foi que realmente houve uma ativação consistente da enzima xantina oxidase, indicando uma situação catabólica do metabolismo de ATP. O decréscimo dos níveis de ATP em combinação com o acúmulo de hipoxantina e ácido úrico no plasma, em decorrência de exercício exaustivo, poderia implicar que o tecido muscular estaria submetido a um nível de estresse metabólico similar ao observado em casos patológicos de esquemia (Sjödin et al., 1990; Sjödin, Westing, 1990).
Os Íons amônio produzidos pela incapacidade do músculo em refosforilar adenosina difosfato (ADP) em ATP representam um fator limitante no prolongamento dos eventos de contração muscular, por atuarem modificando a atividade neuromuscular e, desta forma, contribuindo para a instalação da fadiga muscular (fadiga periférica) (Cairns, 2006). Além disto, a amônia, ao atingir o sistema nervoso central – o que induz efeito supressor funcional (fadiga central) - conduz a diminuição de recrutamento de unidades motoras (Mutch, Banisteer, 1983). A formação da amônia ocorre pelo ciclo nucleotídeo adenina (ADP → AMP + H2O → IMP + NH3). Este ciclo é ativo também no cérebro e em outros órgãos (Urhausen, Kindermann, 1992); a partir deste processo ocorre elevação nos níveis de amônia sanguínea (Harris, Dudley, 1989), sendo que a taxa e período de acúmulo de amônia dependem do tipo de exercício e de sua duração (Itoh, Ohkuwa, 1990). Experimentos sobre o efeito de exercícios de endurance em cicloergômetro em intensidades moderada e alta observaram que, com o aumento da intensidade e duração do exercício, a produção de amônia pelo músculo-esquelético também aumentava devido à incapacidade muscular de refosforilar ADP, ocorrendo redução no recrutamento de unidades motoras. Bangsbo e colaboradores (1996) observaram que durante a execução de exercícios intermitentes para braços e pernas em homens jovens e ativos, nas intensidades média e alta, ocorreu aumento na concentração de amônia muscular liberada para o sangue. No descanso, as concentrações de amônia no sangue arterial foram reduzidas, retornando ao músculo, possivelmente sendo utilizada para a formação de uréia e sendo excretada.
As análises das concentrações plasmáticas de creatinina e uréia permitiram estabelecer uma visão mais integrada do estresse metabólico induzido pelo treinamento contínuo. A uréia, em condições metabólicas normais, é um produto final da degradação dos aminoácidos excedentes, haja vista que os seres vivos não são capazes de armazenar aminoácidos nem proteínas. A degradação do aminoácido ocorre por meio da remoção e excreção do grupo amino e a oxidação da cadeia carbônica remanescente. O grupo amino removido é convertido a uréia e as cadeias carbônicas são convertidas a compostos comuns ao metabolismo de carboidratos e lipídios. Apesar deste parâmetro ser apresentado como um índice sintomatológico da síndrome do overtraining, resultados salientam que o exercício físico possui um efeito catabólico sobre o metabolismo protéico acompanhado simultaneamente por respostas anabólicas (Smith, 2000). O turnover (relação síntese-degradação) protéico é uma resposta regulatória à sobrecarga do treinamento físico. Podemos observar que, a partir da análise da quarta semana, todas as outras análises de concentração plasmática de uréia permaceram num mesmo patamar de valor de mediana e dispersão dos quartis, sugerindo que esse parâmetro metabólico não pode, por si só, ser considerado um indicador seguro de alteração metabólica.
A creatinina é um produto da degradação da fosfocreatina. O sistema ATP-Fosfocreatina constitui um sistema imediato de energia para a contração muscular de rápida duração e alta intensidade, sendo um processo estritamente anaeróbio. A creatinina é liberada na circulação e excretada pela urina. A quantidade excretada diariamente é constante, pelo fato de sua formação ser proporcional à massa muscular. Desta forma, ela torna-se um sensível indicador de função renal e alteração metabólica muscular. Nossos resultados mostram uma significativa queda na concentração plasmática de creatinina na análise da quarta semana, a qual coincide com o momento pico de sobrecarga de corrida. Tal como visto para os valores de uréia, era de se esperar que, caso houvesse alguma alteração deste parâmetro, o aumento progressivo de intensidade do treinamento pudesse, ao invés da diminuição, elevar os índices de creatinina. A literatura não aponta evidências para esta adaptação. Uma possível interpretação seria um aumento na filtração de creatinina pelos rins, em virtude de alguma alteração hormonal. Isto poderia ser comprovado pela dosagem de creatinina excretada pela urina, condição não analisada neste trabalho. A análise da semana 8 apresentou um quadro bem homogêneo deste parâmetro metabólico, tal como visto para o grupo controle. As demais análises (2x/dia, 3x/dia e 4x/dia) permaneceram num mesmo patamar, com grande dispersão dos resultados, o que dificulta estabelecer uma relação direta com os demais marcadores estudados.
Concluindo, podemos considerar que o estresse metabólico induzido pelo protocolo de indução de overtraining gerou uma condição diversificada de respostas fisiológicas. Os marcadores de catabolismo muscular (uréia e creatinina) não sofreram alterações sensíveis ao longo de todo o processo de treinamento contínuo, indicando que alterações bioquímicas – sobretudo aquelas relacionadas aos níveis de estresse oxidativo (Antunes Neto et al., 2006) precedem os eventos iniciais de instalação do overtraining. A análise isolada dos valores para ácido úrico, que apresentaram aumento significativo no momento de maior sobreexigência orgânica, estabelece apenas um parâmetro de indicação de alteração fisiológico para um processo considerado multivaríavel. Portanto, torna-se desejável que novos procedimentos protocolares sejam desenvolvidos com a utilização de animais, com o intuito de compreender relações mais explícitas sobre o fenômeno complexo do overtraining.
Referências bibliográficas
-
ANTUNES NETO, J. M. F.; TOYAMA, M. H.; CARNEIRO, E. M.; BOSCHERO, A. C.; PEREIRA-DA-SILVA, L.; MACEDO, D. V. Circulating leukocyte heat shock protein 70(hsp70) and oxidative stress markers in rats after a bout of exhaustive exercise. Stress, v. 9, n. 2, p. 107-115, 2006.
-
ANTUNES NETO, J. M. F.; PEREIRA-DA-SILVA, L.; MACEDO, D. V. Heat shock proteins as oxidative stress markers in rats submitted to exhaustive intermittent running training. Brazilian Journal of Biomotricity, v. 2, n. 3, p. 160-175, 2008.
-
ANTUNES NETO, J. M. F.; PEREIRA-DA-SILVA, L.; MACEDO, D. V. Proteínas de estresse “HSP70” atuam como marcadoras de estresse oxidativo em ratos “Wistar” submetidos a treinamento contínuo de corrida para indução de “overreaching”. Revista Brasileira de Educação Física, Esporte, Lazer e Dança, v. 3, n. 4, p. 116-130, 2008.
-
BANGSBO, J.; KIENS, B.; RICHTER, A. Ammonia uptake in inactive muscles during exercise in humans. American Journal of Physiology, Endocrinology and Metabolism, v. 270, n. 1, p. E101-E106, 1996.
-
BOMPA, T. O. Theory and methodology of training: to key to athletic performance. Dubuque: Kendall/Hunt, 1990.
-
CAIRNS, S. P. Lactic Acid and Exercise Performance: Culprit or Friend? Sports Medicine, v. 36, n. 4, p. 279-291, 2006.
-
CÓRDOVA, A.; NAVAS, F. J. Os radicais livres e o dano muscular causado pelo exercício: papel dos antioxidantes. Revista Brasileira de Medicina do Esporte, v. 6, n. 5, p. 204-208, 2000.
-
EICHNER, E. R. Overtraining: consequences and prevention. Journal of Sports Sciences, v. 13, p. S41-S48, 1995.
-
HARRIS, R. T.; DUDLEY, G. A. Exercise alters the distribution of ammonia and lactate in blood. Journal of applied Physiology, v. 66, n. 1, p. 313-317, 1989.
-
ITOH, H.; OHKUWA, T. Peak blood ammonia and lactate after submaximal, maximal and supramaximal exercise in sprinters and long-distance runners. European Journal of Applied Physiology and Occupational Physiology, v. 60, n. 4, p. 271-276, 1990.
-
JOHNSON, M. B.; THIESE, S. M. A review of overtraining syndrome: recognizing the signs and symptoms. Journal of Athletic Training, v. 27, n. 04, p. 352-354, 1992.
-
KIBLER, W. B.; CHANDLER, T. J.; STRACENER, E. S. Musculokeletal adaptations and injuries due to overtraining. Exercise and Sports Sciences Reviews, v. 20, p. 99-127, 1992.
-
KUIPERS, H. How much is too much? Performance aspects of overtraining. Research Quarterly for Exercise and Sport, v. 67, n. 03, p. S65-S69, 1996.
-
KUIPERS, H.; KEIZER, H. A. Overtraining in elite athletes: review and directions for the future. Sports Medicine, v. 6, n. 4, p. 79-92, 1988.
-
LAPIM, L. P.; PRESTES, J.; PEREIRA, G. B.; PALANCH, A. C.; CAVAGLIERI, C. R.; VERLENGIA, R. Respostas metabólicas e hormonais ao treinamento físico. Revista Brasileira de Educação Física, Esporte, Lazer e Dança, v. 2, n. 4, p. 115-124, 2007.
-
LEHMANN, M.; FOSTER, C.; DICKHUTH, H. H.; GASTMANN, U. Autonomic imbalance hypothesis and overtraining syndrome. Medicine and Science in Sports and Exercise, v. 30, n. 7, p. 1140-1148, 1998.
-
LEHMANN, M., FOSTER, C., KEUL, J. Overtraining in endurance athletes: a brief review. Medicine and Science in Sports and Exercise, v. 25, n. 07, p. 854-862, 1993.
-
MCGILL, R.; TUKEY, J. W.; LARSEN, W. A. Variations of Box Plots. American Statistician, v. 32, n. 01, p. 12-16, 1978.
-
MUTCH, B. J.; BANISTEER, E. W. Ammonia metabolism in exercise and fatigue: a review. Medicine and Science in Sports and Exercise, v. 15, n. 1, p. 41-50, 1983.
-
PASCOE, D. D.; COSTILL, D. L.; FINK, W. J. Glycogen resynthesis in skeletal muscle following resistive exercise. Medicine and Science in Sports and Exercise, v. 25, n. 03, p. 349-354, 1993.
-
SJÖDIN, B.; WESLING, H.; APPLE, S. Biochemical mechanisms for oxygen free radical formation during exercise. Sports Medicine, v. 10, p. 236-254, 1990.
-
SJÖDIN, B.; WESTING, Y. H. Changes in plasma concentration of hypoxanthine and uric acid in man with short-distance running at various intensities. International Journal of Sports and Medicine, v. 11, p. 493-495, 1990.
-
SMITH, L. L. Cytokine hypothesis of overtraining: a physiological adaptation to excessive stress? Medicine and Science in Sports and Exercise, v. 32, n. 02, p. 317-331, 2000.
-
TENENBAUM, G. Theoretical and practical considerations in investigating motivation and discomfort during prolonged exercise. Journal of Sports Medicine and Physical Fitness, v. 36, n. 03, p. 145-154, 1996.
-
TIIDUS, P. M. Radical species in inflamation and overtraining. Canadian Journal of Physiology and Pharmacology, v. 76, n. 5, p. 533-538, 1998.
-
TRAVERSO, A.; OTTONELLO, G. A.; SENAREGA, D. Exercice physique et prévention de la douleur musculaire squelettique due à des facteurs psychologiques. Utilité diagnostique du EMG-biofeedback. Médecine du Sport, v. 70, n. 05, p. 223-230, 1996.
-
URHAUSEN, A.; KINDERMANN, W. Blood Ammonia and lactate concentrations during endurance exercise of differing intensities. European Journal of Applied Physiology, v. 65, n. 3, p. 209- 214, 1992.
Outros artigos em Portugués
| |

Búsqueda personalizada
|
|---|---|
|
EFDeportes.com, Revista Digital · Año 17 · N° 170 | Buenos Aires,
Julio de 2012 |
|