
|
|||
|---|---|---|---|
|
|
Effects of upper body exercise on knee joint position sense |
|
|
|
*Universidad Pablo de Olavide Sevilla **EADE (Málaga). University of Wales. (España) |
Alejandro Camps Olmedo* Juan Rojo Rodríguez** Miguel Angel García de la Concepción** |
|
|
|
Abstract The aim of this paper was to determine the immediate effects of an upper body fatigue protocol on knee joint position sense, measured by a position matching technique, we measured three angles, 16º-32º-48º. 20 fatigue subjects and 20 control subjects participated in this study. Using the Borg scale, we also measured ratings of perceived exertion before, during, and after the fatigue protocol or rest period. We found a significant increase in Absolute Angle Error (AAE) for the three angles from pretest to posttest in the fatigue group (16º= 7.2º ± 4.1º vs 10.8º ± 6.5º, 32º = 6.2º ± 3.8º vs 10.7º ± 6.3º, 48º = 4.3º ± 3.3º vs 5.6º ± 3.3º ), no significant differences from pretest to posttest in the control group for the three angles (16º= 7.1º ± 3.7 º vs 8.2º ± 3.8º, 32º = 5.3º ± 3.6º vs 5.1 ± 3.2º, 48º = 4.3º ± 2.7º vs 4.2º ± 3.1º ), no significant difference between control and fatigue group in pretest (16º = 7.1º ± 3.6 º vs 7.2º ± 3.9º, 32º = 5.3º ± 3.6 º vs 6.2º ± 3.8 º, 48º = 4.2º ± 2.7º vs 4.3º ± 3.3º ), and a significant difference between control and fatigue group on the posttest (16º = 8.2º ± 3.8º vs 10.8º ± 6.5º, 32º = 5.1º ± 3.2º vs 10.7º ± 6.3º, 48º = 4.2º ± 3.1º vs 5.6º ± 3.3º ). The rating of perceived exertion scores were significantly different between the fatigue and control group at the middle (10.5 ± 1.1 vs 2.5 ± 0.6) and end (19.3 ± 0.7 vs 2.5 ± 0.6) of the fatigue or rest period. Our results demonstrated that the AAE increased immediately after the fatigue protocol, demonstrating that joint position sense of the knee diminished. Since our subjects performed an upper body fatigue protocol and this muscular group do not play any role in joint position sense of the knee we can ensure that the decrease in joint position sense was due to central fatigue. Keywords: Joint position sense. Fatigue. Surfboard riding. Upper body exercise. |
|||
|
|
http://www.efdeportes.com/ Revista Digital - Buenos Aires - Año 14 - Nº 136 - Septiembre de 2009 |
|
|
1 / 1
Introduction
Throughout the history of the sport and physical activity, it has been attempted to improve our physical qualities that enable us to delay fatigue. These adaptations would make it possible for us to maintain a physical activity through time and, in addition, to perform precise and controlled movements.
Muscle fatigue is a complex phenomenon that has been defined as a reduction in the force-generating capacity regardless of the task performed (Bigland-Ritchie and Woods 1984).
Excessive workloads can increase physical fatigue, resulting in diminished physical endurance and performance and, ultimately, a reduction in productivity. In addition, a combination of muscular fatigue and general subjective fatigue may also manifest itself by causing temporary interference with the functioning of the central nervous system (CNS) as well as the peripheral nervous system (Gandevia 1998; Lepers et al. 2002). It is known that muscular fatigue modifies both the peripheral proprioceptive system but also the central processing of proprioception (Forestier et al. 2002; Sharpe and Miles 1993). More specifically, during local muscle fatigue, nociceptors are activated by metabolic products of muscular contraction including bradykinin (Mense and Meyer 1988), arachidonic acid and prostaglandin E2 (Rotto and Kufman 1988), potassium (Rybicki et al. 1985), and lactic acid (Rotto and Kufman 1988). These metabolites and inflammatory substances have a direct impact on the discharge pattern of muscle spindles (Pedersen et al 1998) that represent the peripheral component of fatigue. In addition, it has been shown that efferent and afferent neuromuscular pathways are modulated via reflexes originating from small-diameter muscle afferents (namely group III and IV afferents) (Bigland-Ritchie et al. 1986; Gandevia 1998). These reflexes originating from the muscle and generated in response to metabolic changes that accompany fatigue could modify the central processing of proprioception that represent the central component of fatigue. Motor cortical and supraspinal factors must also be considered in case of muscle fatigue. More precisely, in fatiguing exercise, it has been shown that cortico-motor neuronal cells firing rate decrease and motor-evoked potentials (assessed with transcranial magnetic stimulation) increase. These results emphasize an inadequate cortical output with fatigue (Gandevia 1998).
Currently the consensus is that proprioception is a complex entity encompassing several different submodalities, for example the sensation of position, velocity, movement detection and force (Gandevia and Burke 1992). Position sense can be functionally defined as the awareness of the actual position of the limb. A few studies have shown that high-intensity exercise to fatigue significantly reduces position sense acuity (Lattanzio et al. 1997; Skinner et al. 1986; Voight et al. 1996). Other studies using this mode of activity, however, found no fatigue-related changes in position sense (Marks and Quinney 1993; Sharpe and Miles 1993; Sterner et al. 1998). To date, no study has examined the effects on position sense of the knee due to fatigue of the upper extremities.
One problem with the current research is that it often uses a repeated isokinetic exercise, or an isometric hold to fatigue the muscle or muscle group, these activities are usually not functional or sport related in anyway. Research needs to support a more functional or sport-related fatiguing protocol to observe how rigors of surfboard paddling performed previous riding the wave in surfboarding affects the position sense of the knee during the wave riding. We wondered what would happens when an exercise involving an upper body muscular group is done and subsequently, performance with a lower body muscular group is sought, that is, if the fatigue of the upper body muscular group affects the position sense of the knee.
One example is surfboarding, in which competitors must row and transport the board out to sea by using their upper extremity muscles. However, the competition only measures the ride of the wave involving all the lower muscles.
Therefore the purpose of this study is to examine how joint position sense of the knee as measured by a position matching technique, is affected by an activity related fatiguing protocol performed with the upper body and simulating a surfboard paddling.
Material and methods
Subjects
Forty male sports sciences students (age= 20.6 ± 1.9 years, Height = 175.6 ± 9.7 cm, Mass = 75.5 ± 16.1 kg) were tested twice during 1 session. Any subject who had suffered a musculoskeletal injury to a lower extremity or a head injury in the 6 months before testing was excluded from the study. We screened subjects for any preexisting visual, vestibular, or balance disorders through self-report. Subjects were randomly assigned to one of the two test groups, control or fatigue. All subjects read and signed the informed consent form approved by the institutional review board, which also approved the study procedures.
Testing apparatus
A purpose built padded frame permitted the leg under examination to rest in a totally relaxed manner (Fig. 1). An electrogoniometer (Penny and Giles Blackwood Ltd, Blackwood, Gwent, Wales, UK) tested to be accurate to 0.5º, was attached to the thigh and shin using padded strapping. The subject sat on a cube with one leg supported on the frame, with a screen preventing any visual clues as to the position of the leg.
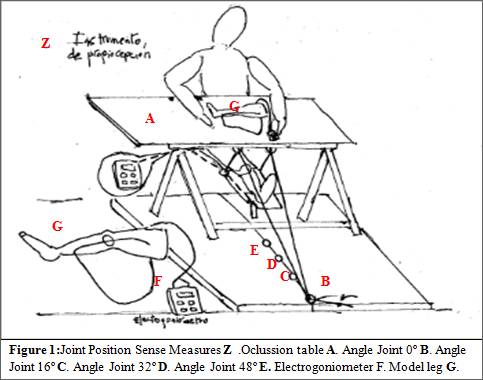
The test as previously describe (Warren et al. 1993), involved the passive movement of the leg to a predetermined sequence of three positions of knee flexion between 0 and 60º. The experimenter placed the leg at one of three test positions; 16º, 32º or 48º from horizontal (Djupsjöbacka and Domkin 2005). Once the leg had been positioned at the test angle, the blindfolded subject was asked reproduce the angle in a held model leg. The subjects indicated verbally when they thought the legs were matched. Position errors were calculated as the difference in angle between the two legs. Each measurement was repeated three times, presenting different angles in random order. The same procedure was used in all subsequent measurements. For the fatigue experiments, measurements were made immediately before and after the exercise. The output from the electrogoniometers was fed into a computer.
Fatigue protocol
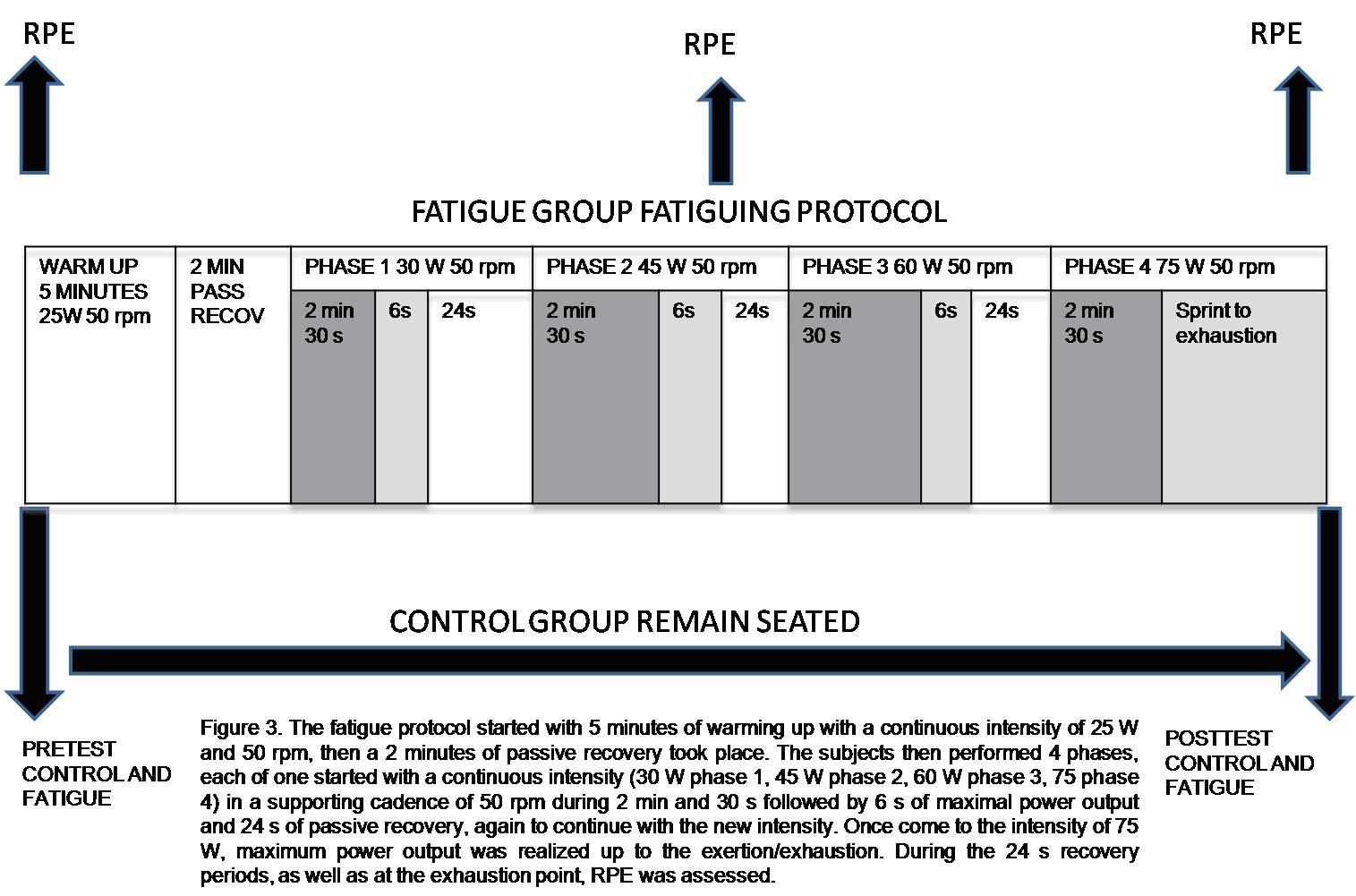
After the familiarization period, all subjects underwent a pretest in which they performed 3 trials. Subjects in the control group then rested, as long as the duration of the fatigue protocol (15.3 ± 2.2 min), whereas subjects assigned to the fatigue group performed the fatigue protocol, which was immediately followed by posttest. The fatigue protocol was performed lying prone on a table (height 40 cm, wide 30 cm, long 130 cm) and using an arm crank ergometer (Monark 881E, Monark-crecent AB, Varberg, Sweden) (Fig. 2).

The fatigue protocol started with 5 minutes of warming up with a continuous intensity of 25 W and 50 rpm, then a 2 minutes of passive recovery took place. The subjects then performed 4 phases, each of one started with a continuous intensity (30 W phase 1, 45 W phase 2, 60 W phase 3, 75 phase 4) in a supporting cadence of 50 rpm during 2 min and 30 s followed by 6 s of maximal power output and 24 s of passive recovery, again to continue with the new intensity. Once come to the intensity of 75 W, maximum power output was realized up to the exertion/exhaustion. Before protocol started, during the 24 s recovery period in phase 2, as well as at the exhaustion point, RPE was assessed. It is important to realize that not every subject was able to reach phase 4, but we must be sure every subject quit at his/her point of exertion/exhaustion or reaching RPE levels up to 15 in the 15-point Borg scale (6-20) (Borg 1970) (Fig. 3).
RPE
We used a 15-point Borg scale (6-20) (Borg 1970) to measure ratings of perceived exertion (RPE) in attempt to quantify the amount of exertion displayed by each subject before, midway through, and after the exertion protocol. The 15-point Borg scale has been recommended for applied investigators of perceived exertion and for predictions of exercise intensity during sports (Borg 1982).
Statistical analysis
Using SPSS software (version 14.0; SPSS inc, Chicago, IL), a series of 3 independent t test was used to examine the differences in absolute angles errors in the three angles as a function of group (control and fatigue), in pretest and posttest. A separate independent t test was used to evaluate RPE. Dependent t tests were used to evaluate the differences between pretest and posttest separately in each group as well as for evaluation of RPE differences in pretest and posttest in each group. We set our alpha level priori at .05.
Results
No significant differences (P > .05) in absolute angles errors were found in the 3 angles between control and fatigue group in the pretest (16º = 7.1º ± 3.6 º vs 7.2º ± 3.9º, 32º = 5.3º ± 3.6 º vs 6.2º ± 3.8 º, 48º = 4.2º ± 2.7º vs 4.3º ± 3.3º ). The opposite case was shown in the posttest where we found significant differences (P < .05) in absolute angles errors in the 3 angles between control and fatigue group (16º = 8.2º ± 3.8º vs 10.8º ± 6.5º, 32º = 5.1º ± 3.2º vs 10.7º ± 6.3º, 48º = 4.2º ± 3.1º vs 5.6º ± 3.3º ) (Fig. 4).
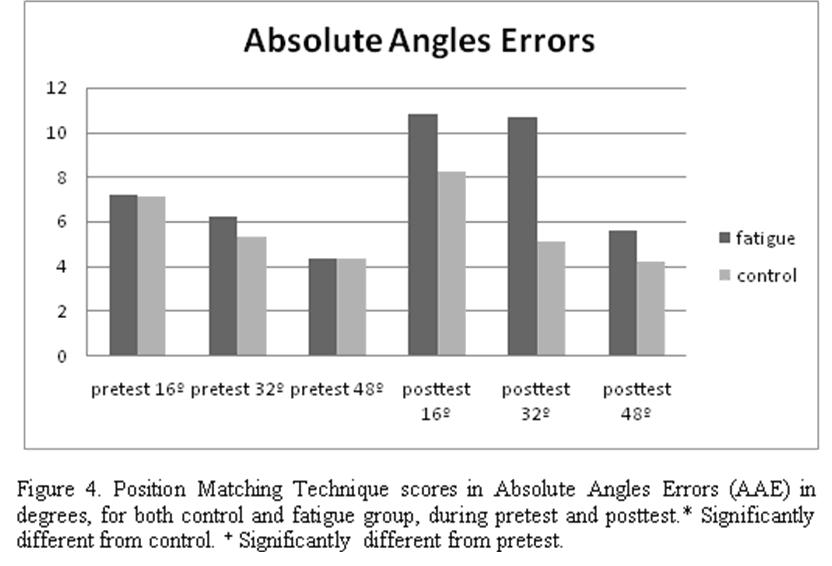
Whereas no significant differences (P > .05) were found between pretest and posttest in control group in any of the 3 angles (16º= 7.1º ± 3.7 º vs 8.2º ± 3.8º, 32º = 5.3º ± 3.6º vs 5.1 ± 3.2º, 48º = 4.3º ± 2.7º vs 4.2º ± 3.1º), the opposite case was shown in the fatigue group, showing the 3 angles significant differences (P < .05) between pretests and posttest ( 16º= 7.2º ± 4.1º vs 10.8º ± 6.5º, 32º = 6.2º ± 3.8º vs 10.7º ± 6.3º, 48º = 4.3º ± 3.3º vs 5.6º ± 3.3º ). This data confirm an increase in absolute angles errors that demonstrate a decreased in joint position sense of the knee after fatigue (Fig. 5, Table 1).
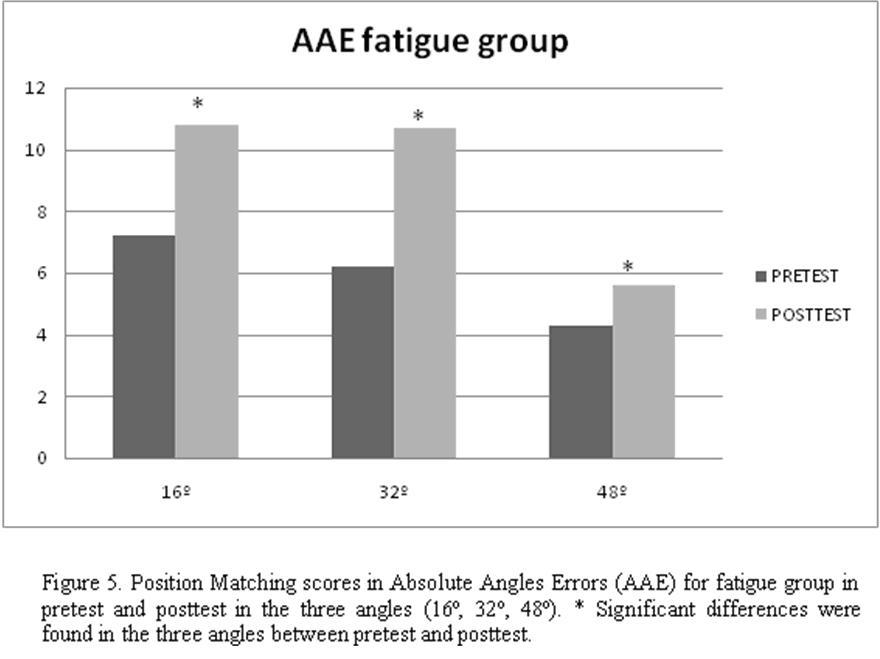
Table 1. Absolute Angles Errors (AAE) for the three angles (16º,32º and 48º) for both control and fatigue group in pretest and posttest
|
Angles |
Control (n = 20) |
Fatigue (n = 20) |
||
|
|
Pretest |
Posttest |
Pretest |
Posttest |
|
16º |
7.1 ± 3.7º |
8.2 ± 3.8º |
7.2 ± 4.1º |
10.8 ± 6.5º* |
|
32º |
5.3 ± 3.6º |
5.1 ± 3.2º |
6.2 ± 3.8º |
10.7 ± 6.3º* |
|
48º |
4.3 ± 2.7º |
4.2 ± 3.1º |
4.3 ± 3.3º |
5.6 ± 3.3º* |
Mean ± SD of AAE in degrees for the three angles (16º, 32º, 48º).
*Statistically significant (P < 0.05) from pretest.
The RPE scores were significantly different (P < .05) between the fatigue and control group at mid part (phase 3) (2.5 ± .6 vs 10.5 ± 1.1) and end ( 2.5 ± .6 vs 19.3 ± .7). The fatigue group demonstrated increased RPE scores at every point. No differences (P > .05) between groups were found in the RPE scores taken at the beginning of the fatigue or rest period (2.5 ± .6 vs 2.6 ± .6) (Fig. 6, Table 2).
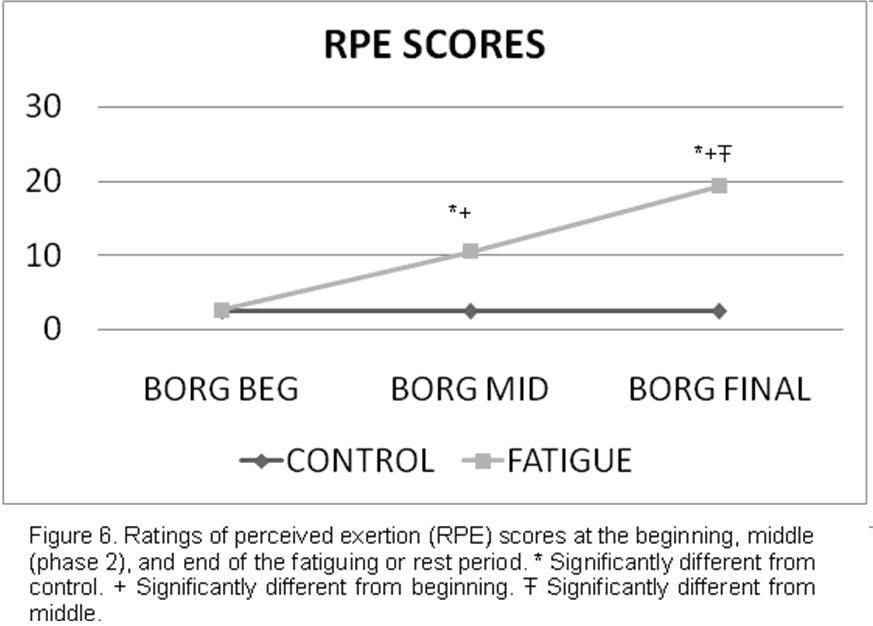
Table 2. Ratings of Perceived Exertion at the Beginning, Middle (Phase 2), and End of the Fatigue or Rest Period.
|
|
Control (n=20) |
Fatigue (N=20) |
||||
|
|
BEG |
MID |
END |
BEG |
MID |
END |
|
Range |
2 |
2 |
2 |
2 |
3 |
2 |
|
Minimum |
2 |
2 |
2 |
2 |
9 |
18 |
|
Maximum |
4 |
4 |
4 |
4 |
12 |
20 |
|
Mean |
2.50 |
2.50 |
2.50 |
2.65 |
10.50* |
19.30* |
|
SD |
.60 |
.60 |
.60 |
.67 |
1.10 |
.73 |
*Statistically significant (P < 0.05) from beginning.
Discussion
Our main findings were that the fatigue group increased significantly absolute angles errors on the posttest than on the pretest and increased significantly absolute angles errors compared with control group in the posttest. Our overall findings demonstrated a decrease in joint position sense of the knee as a result of fatigue as measured by Position Matching Technique.
Our protocol was designed to replicate the fatigue surfboard riders would experience during the course of a paddling action used to pass over the wave breaking section and then “catch” the wave. We feel confident that the changes in RPE scores demonstrated that our subjects were fatigued to a level representative of working at 100% of maximal heart rate or maximal oxygen uptake (VO2max) and that this fatigue elicited joint position sense during posttest.
Ratings of perceived exertion
One potential limitation to studies of central fatigue is the ability to quantify the amount of fatigue to which the subjects are subjected. It would be difficult to compare the effect of fatigue on joint position sense if the degree of fatigue varied across investigations. To rectify this, several groups, including ours, have used the Borg RPE scale in an attempt to quantify the amount of fatigue (Nardone et al. 1997; Robertson et al. 2000; Seliga et al. 1991; Wilkins et al. 2004).
Because perceived exertion and VO2max are highly correlated, the RPE scale may be used as a substitute to determine exercise intensity (Brooks et al. 1996). Nardone et al. (1998) employed the 10-point Borg RPE scale and found that their protocol elicited perceived exertion that was classified between 5 and 7 (strong to very strong). Using the 15-point Borg scale, Seliga et al. (1991) showed perceived exertion scores increased significantly with increase in work load. The RPE values ranged from 9 to 10 during a light workload to 11 to 12 during a moderate workload to 14 to 16 during a heavy workload. They also noted that sway values were higher after exercise at higher workloads.
Other investigators have correlated RPE with percentage of maximum heart rate reserve, percentage of ventilator threshold, or percentage of VO2max during various exercise tasks (Mahon et al. 1997; Robertson et al. 2000). In a group of physically active males, RPE was measured during treadmill exercise, cycle exercise, and simulated ski exercise at 70%,80% and 90% VO2max across the exercise modes to 15.4 to 16 at 80% and 18 to 18.2 at 90% (Robertson et al. 2000).
Similarly, RPE has been recorded during 10-minute graded cycle-ergometer testing in males (25.3 ± 2.0 years) in relation to ventilator threshold (Mahon et al. 1997), the RPE values were 10.2 ± 1.2 at the 5-minute mark of an exercise at 80% ventilator threshold (heart rate = 124.6 ± 14.3 beats/minute) and 15.8 ± 1.7 at the 10-minute mark of exercise at 120% ventilator threshold (heart rate = 168.9 ± 13.5 beats/minute) (Mahon et al. 1997).
The RPE values noted in afore mentioned investigations are lower than our findings of 10.5 ± 1.1 and 19.3 ± .7 during the middle (phase 2) and end of our fatigue protocol, respectively.
Therefore, we feel confident that, during our protocol, subjects were working at a level greater than 90% VO2max at the end of protocol, and the decrease in position matching performance noted during the posttest was a result of their fatigue.
Fatigue and Joint Position Sense
The present study demonstrates clearly that position sense acuity decreased (i.e. increase in absolute error) following a fatiguing protocol. Deficits in position sense acuity following muscle fatigue have been demonstrated in previous studies for the knee (Skinner et al. 1986; Lattanzio et al. 1997) and for the shoulder joints (Voight et al. 1996), which are in general agreement with our findings. However, other studies for the knee (Marks and Quinney 1993), elbow (Sharpe and Miles 1993) and shoulder joint (Sterner et al. 1998) revealed no such effects. For nearly all of the previous reports, fatigue was elicited by imposing high-intensity isometric or isokinetic exercise protocols, and fatigue was assessed as a relative change in muscle force. Other study (Björklund et al. 2000), focused on the shoulder due to the high incidence of neck-shoulder work-related problems, examined and showed a decreased in position sense following low-intensity repetitive work to fatigue. But no study has investigated the effects of an upper body exercise on knee joint position sense. Position sense acuity is been believed to be to a large extent dependent upon muscle spindle activity, the effect that fatigue has on the muscle spindle system is been usually the plausible mechanism for most of previous findings. A direct impact of fatigue on the discharge patterns of muscle spindles was demonstrated in an animal study (Pedersen et al. 1998). This finding of muscle spindle alterations was extended to show that the information transmitted by ensembles of primary afferents is reduced following fatigue. Fatigue may therefore result in a reduction in position sense acuity. A further detailing of the mechanism may reside in the accumulation of metabolites and/or inflammatory substances within the muscle during activity to fatigue (i.e. lactic acid or bradykinin). Both metabolites and inflammatory substances have been shown to elicit increased muscle spindle static and/or dynamic sensitivity via reflex-mediated pathways from chemosensitive group III and IV afferents ontoc-motoneurones (Djupsjöbacka et al. 1994, 1995; Johansson et al. 1993; Pedersen et al. 1997).
In opposite of these findings in our study we performed an exercise of upper body and then we examined its effects in the lower body, so it would be difficult that the prejudices produced in the position sense of knee were due to a peripheral fatigue, in this case of the peripheral receptors (principally the SP) and its afferences Ia and II. Therefore, it is necessary to reject the possibility of the peripheral fatigue, so we establish a relation between central fatigue, central governor theory (Noakes et al. 2005) and sense of effort as responsible for the decrease in joint position sense.
Some authors show conclusions, (Carrasco et al. 2005) that make clear that the effects obtained on the proprioception depend on the type of test and the effect that this one produces on the fatigue besides the protocol used, coming to the conclusion that the aptitude to reproduce a certain joint position in situation of fatigue, is not due to a loss of peripheral signs, but to a deficient processing of the information to central level. Other authors (Allen and Proske 2006; Proske 2004; Walsh et al. 2005;); showed that the joint position sense uses the sensation of the effort based on the effects of the muscular fatigue and the muscular damage. Departing from an absence of fatigue also other authors (Ansems et al. 2006; Semmler and Miles in their review of 2006 (mentioning Gandevia, 1996)) showed that the proprioception does not depend only of the receptors but it is centrally generated as well. These central generated signs are associate with the sensation of effort, which are produced for motor efferent that need the accomplishment of voluntary contractions. The peripheral afferent are at the time compared with the sensation of effort to provide the perception of the relative position of our member. (Gandevia 1996; mentioned by Semmler and Miles, 2006). All our assumptions agree with Gandevia (1996) about the sense of effort, since it connects with the feeling of fatigue.
All this findings agree with the theories of Noakes et al. (2005) about Central Governor, they conclude that the human body functions as a complex system during exercise. Using feed forward control and in response to afferent feedback from multiple central and peripheral sensors, the brain paces the body during exercise specifically to ensure that the preplanned activity is completed without any loss of cellular homoeostasis. Control is applied by changes in the extent of skeletal muscle recruitment during exercise and by the inhibitory effects of the increasingly disagreeable sensations of fatigue that are generated by the brain during exercise.
The prediction of the central governor model, is that the brain does not recruit additional motor units during prolonged exercise because such additional recruitment would threaten the capacity to maintain homoeostasis, potentially leading to, at best, the premature termination of exercise, or, at worst, organ damage, organ failure, and even perhaps death. This model also predicts the rising perception of discomfort produced by exhausting exercise, the prediction of the central governor model is that, as exercise performance is centrally regulated by the CNS, then fatigue should no longer be considered a physical event but rather a sensation or emotion, separate from an overt physical manifestation, and peripheral metabolites have signaling properties that assist in determining the continuously reset pacing strategy, and are not merely energy substrates or inert metabolic byproducts. As argued by Lambert et al. (2005) the natural conclusion of each researcher examining any single metabolite is that this specific metabolite is the ultimate regulator of the complete system.
So changes in any physiological variable results from alterations in neural command or peripheral regulatory systems in response to prior system perturbation. The continuous interaction between feed forward and feedback control mechanisms in the brain and peripheral physiological systems produces a robust, self-sustaining mechanism that maintains homoeostasis by ensuring that no system is ever overwhelmed or used to absolute maximal capacity.
For the above mentioned we can resume that the factors which could potentially cause a decrease in joint position sense after fatigue focus on both central and local means of fatigue. Central or whole-body fatigue, refers to a decrease in the central nervous system output to the muscles (Brooks et al. 1996) and likely has a component that includes factors responsible for the sense of effort in addition to the alterations in motor pathways (McComas 1996). Localized muscle fatigue is induced by a decrease in the metabolic substrates available for muscle contraction, such as adenosine triphosfate, creatine phosphate, and glycogen, as well as an increase in metabolites, including lactic acid, in the muscle, resulting in an inability to maintain a desired muscular force output (Rozzi et al. 2000). Since our subjects performed an upper body fatigue protocol and this muscular group do not play any role in joint position sense of the knee we can ensure that the decrease in joint position sense was due to central fatigue. More in deep we agree with the conclusions shown by Abbiss et al. (2005) about models of fatigue where they argue that the increase in recruitment produce and increase in the sense of effort that produce an increase in central neural activation to produce fatigue as a part of the body’s system of defense, one possible explanation could be therefore, that our subjects suffered high levels of central neural activation as a consequence of the exhaustion achieved at the end of the fatigue protocol, this central neural activation would lead to an increase in the sense of effort that would lead to a deficient processing of the information to central level, and finally, this deficient processing of the information to central level would lead to a decrease in joint position sense at the knee as measured by the position matching technique.
Our findings show an increase in absolute angle errors in position matching technique immediately after the fatigue protocol, demonstrating that joint position sense diminished. The surfboard riders should improve their paddling capacity in order to avoid or delay fatigue during wave riding. Future investigators should find a more surfboard-related test to investigate if the decrease in joint position sense really affects the wave riding itself. In the other hand it would be good to use surfboard riders as subjects, since they must be used to the paddling exercise and the effects may be different. Our findings suggest that during surfboard training, surfboard riders should pay more attention of paddling performance in order to avoid the effects of fatigue achieved during the paddling action during the wave riding. We suggest that trainers should include in their programs some more paddling related exercise such as swimming or rowing as well as paddling muscles specific strength exercises.
Acknowledgments
We thank Ramón Alvero and all his team for the technical assistance and Manuel Rosety for his active support throughout the entire project.
References
-
Abbis CR, Laursen PB (2005). Models to explain fatigue during prolonged endurance cycling. Sports Med 35 (10): 865-898.
-
Allen TJ, Proske U (2006). Effect of muscle fatigue on the sense of limb position and movement. Exp Brain Res 170: 30-38.
-
Ansems G.E, Allen T.J, Proske U (2006). Position sense at the human forearm in the horizontal plane during loading and vibration of elbow muscles. J Physiol 576: 445-455
-
Bigland-Ritchie B, Woods JJ (1984). Changes in muscle contractile properties and neural control during human muscular fatigue. Muscle Nerve 7: 691-9.
-
Bigland-Ritchie B, Dawson NJ, Johansson RS,Lippold OC (1986). Reflex origin for the slowing of motoneurone firing rates in fatigue of human voluntary contractions. J Physiol 379: 451– 459.
-
Björklund M, Crenshaw AG, Djupsjobacka M, Johansson H ( 2000). Position sense acuity is diminished following repetitive low-intensity work to fatigue in a simulated occupational setting. Eur J Appl Physiol 81: 361-367.
-
Borg G (1970). perceived exertion as an indicator of somatic stress. Scand J Rehabil Med 2: 92-98.
-
Borg G. Phychophysical bases of perceived exertion (1982). Med Sci Sports Exerc 14:377-381.
-
Brooks GA, Fahey TD, White TP (1996). Exercise physiology. Mountain view, CA: Mayfield Publishing Co.
-
Carrasco Páez L, Rodríguez Torralba N, Nadal Soler C (2005). Efectos de la fatiga muscular local sobre la propiocepción en la articulación de la rodilla. Archivos de Medicina del deporte.; 14(3): 144-149
-
Djupsjöbacka M, Johansson H, Bergenheim M (1994). Influuences on the gamma-muscle spindle system from muscle afferents stimulated by increased intramuscular concentrations of arachidonic acid. Brain Res 663: 293±302
-
Djupsjöbacka M, Johansson H, Bergenheim M, Wenngren BI (1995). Influences on the gamma-muscle spindle system from muscle afferents stimulated by increased intramuscular concentrations of bradykinin and 5-HT. Neurosci Res 22: 325±333
-
Djupsjöbacka M, Domkin D (2005). Correlation analysis of propioceptive acuity in ipsilateral position matching and velocity-discrimination. Somatosensory and motor research; 22(1/2): 85-93.
-
Forestier N, Teasdale N, Nougier V (2002). Alteration of the position sense at the ankle induced by muscular fatigue in humans. Med Sci Sport and Exerc 34(1): 117-122.
-
Gandevia SC, Burke D (1992). Does the nervous system depend on kinesthetic information to control natural limb movements. Behav Brain Sci 15: 614-632.
-
Gandevia SC ( 1996). Kinaesthesia: role for afferent signals and motor commands, In: Rowell LB, Shepherd JT, editors. Handbook of physiology, Section 12. Exercise: regulation and integration of multiple systems. New York: Oxford University Press: p 128-172.
-
Gandevia SC (1998). Neural control in human muscle fatigue: changes in muscle afferents, moto neurons and motor cortical drive. Acta Physiol Scand 162: 275-283.
-
Johansson H, Djupsjöbacka M, Sjölander P (1993) Influences on the gammma-muscle spindle system from muscle afferents stimulated by KCl and lactic acid. Neurosci Res 16: 49-57
-
Lambert EV, St Clair Gibson A, Noakes TD (2005). Complex systems model of fatigue: integral control and defense of homeostasis during exercise in humans. Br J Sports Med: 39:52–62.
-
Lattanzio P-J, Petrella RJ, Sproule JR, Fowler PJ (1997). Effects of fatigue on knee proprioception. Clin J Sport Med 7: 22-27.
-
Lepers R, Maffiuletti N, Rochette L, Brugniaux J, Millet F (2002). Neuromuscular fatigue during a long-duration cycling exercise. J Appl Physiol 92: 1487-1493.
-
Mahon AD, Duncan GE, Howe CA, Del Corral P (1997). Blood lactate and perceived exertion relative to ventilatory threshold: boys versus men. Med Sci Sports Exerc 29:1332-1337.
-
Marks R, Quinney HA (1993). Effect of fatiguing maximal isokinetic quadriceps contractions on ability to estimate knee-position. Percept Mot Skills 77: 1195-1202.
-
Mense S, Meyer H (1988). Bradykinin-induced modulation of the response behavior of different types of feline group III and IV muscle receptors. J Physiol 398: 49–63.
-
McComas AJ (1996). Skeletal muscle: Form and function. Champaign, IL:Human Kinetics 230-231.
-
Nardone A, Tarantola J, Galante M, Schieppati M (1997). Fatigue effects on bodybalance. Electroencephalogr Clin Neurophysiol 105: 309-320.
-
Nardone A, Tarantola JC, Galante M, Schieppati M (1998). Time course of stabiometric changes after a strenuous treadmill exercise. Arch Phys Med Rehabil 79: 920-924.
-
Noakes TD, Gibson A St Clair, Lambert EV (2005). From catastrophe to complexity: a novel model of integrative central neural regulation of effort and fatigue during exercise in humans: summary and conclusions. Br J Sports Med;39: 120–124.
-
Pedersen J, Sjölander P, Wenngren BI, Johansson H (1997) Increased intramuscular concentration of bradykinin increases the static fusimotor drive to muscle spindles in neck muscles of the cat. Pain 70: 83-91.
-
Pedersen J, Ljubislavlevic M, Bergenheim M, Johansson H (1998). Alterations in information transmission in ensemble of primary muscle spindle afferents after muscle fatigue in heteronymous muscle. Neuroscience 84:953–959.
-
Proske U (2005). What is the role of muscle receptors in proprioception?. Muscle and Nerve. 31: 780-787.
-
Robertson RJ, Moyna NM, Sward KL, Mikkich NB, Gross FL, Thompson PD (2000). Gender comparison of RPE at absolute and relative physiological criteria. Med Sci Sports Exerc 32: 2120-2129.
-
Rotto DM, Kaufman MP (1988). Effect of metabolic products of muscular contraction on discharge of group III and IV afferents. J Appl Physiol 64: 2306–2313.
-
Rozzi S, Yuktanandana P, Pinciverg D, Lephart SM (2000). Role of fatigue on proprioception and neuromuscular control. In : Lephart SM, Fu FH, Eds. Proprioception and neuromuscular control in joint stability. Champaign, IL: Human Kinetics 375-383.
-
Rybicki KJ, Waldrop TG, Kaufman MP (1985). Increasing gracilis interstitial potassium concentrations stimulates group III and IV afferents. J Appl Physiol 58: 936–941.
-
Seliga R, Bhattacharya A, Succop P, Wixhstrom R, Smith D, Willeke K (1991). Effect of work load and respirator wear on postural stability, heart rate, and perceived exertion. Am Ind Hyg Assoc J 52: 417-422.
-
Semmler JG, Miles T (2006). Exercise, effort, and limb position sense. J Appl Physiol 100: 1099-1100.
-
Sharpe MH, Miles TS (1993). Position sense at the elbow after fatiguing contractions. Ex. Brain Res 94: 179–182.
-
Skinner HB, Wyatt MP, Hodgdon JA, Conard DW, Barrack RL (1986). Effect of fatigue on joint position sense of the knee. J Orthop Res 4: 112-118.
-
Sterner RL, Pincivero DM, Lephart SM (1998). The effects of muscular fatigue on shoulder proprioception. Clin J Sport Med 8: 96-101.
-
Voight ML, Hardin JA, Blackburn TA, Tippett S, Canner GC (1996). The effects of muscle fatigue on and the relationship of arm dominance to shoulder proprioception. J Orthop Sports Phys Ther 23: 348-353.
-
Walsh LD, Hesse CW, Morgan DL, Proske U (2004). Human forearm position sense after fatigue of elbow flexor muscles. J Physiol 558.2: 705-715.
-
Warren PJ, Olanakuhn TK, Cobb AG (1993). Propioception after knee arthroplasty: the influence of prosthetic design. Clin Orthop; 297: 182-7.
-
Wilkins JC, Valovich McLeod TC, Perrin DH, Gansneder BM (2004). Performance on the balance error scoring system decreases after fatigue. J Athl Train 39(2): 156-161.
Another articles in English
| |
|
|---|---|
|
revista
digital · Año 14 · N° 136 | Buenos Aires,
Septiembre de 2009 |
|